АНАЛИЗ ПРИЧИН
ЗАМЕДЛЕНИЯ РОСТА ЖИВЫХ ОРГАНИЗМОВ
УДК 577.95 © 2005 г. Седова Г.П.
В данной статье предпринята
попытка объяснить причину замедления роста
живых организмов.
Предлагаемая статья продолжает тему роста живых организмов. В двух предыдущих статьях «Закономерности роста биологических объектов» («Математическая морфология», 2004 г.) и «Сравнительный анализ эмбрионального и постэмбрионального роста живых организмов» («Математическая морфология», 2005 г.) была установлена закономерность замедления роста живых организмов. Причина, по которой происходит это замедление, в указанных статьях не рассматривалась. Попытаемся теперь подойти к этой причине.
Как и в предыдущих статьях будем сравнивать рост популяции одноклеточных организмов при наличии нормальных условий и рост многоклеточного организма и находить между этими процессами сходство и различие.
Если предположить, что в процессе роста с течением времени какая-то субстанция истощается, то это происходило бы и в популяции одноклеточных и в многоклеточном организме, т.к. процесс роста и в том и другом случае происходит одинаково путем деления клеток и их роста. Но рост популяции одноклеточных организмов при нормальных условиях происходит без замедления, с постоянным периодом удвоения массы, в то время как рост многоклеточного организма резко замедляется в самом начале процесса. Как писал акад. Шмальгаузен И.И., рост цыпленка падает уже после 2-х суток насиживания. В чем же заключается причина этого различия?
В популяции одноклеточных в момент деления материнской клетки дочерние клетки отделяются друг от друга, образуя самостоятельные организмы. Как писал известный исследователь процесса митоза Д. Мэзия, вновь образовавшиеся дочерние клетки, способные подобно амебе к передвижению, почувствовав себя свободными, тот час же пытаются отползти друг от друга.
Клетки же многоклеточного организма разделившись, остаются тем или иным способом связанными между собой, образуя общую массу. Видимо в этом и заключена причина, по которой рост многоклеточного организма замедляется с течением времени.
Как было показано в первой из указанных статей, отдельная особь популяции одноклеточных организмов растет по общему закону, линейно увеличивая период удвоения своей массы. Но в момент разделения массы материнской клетки на две после митоза каждая из дочерних клеток, избавившись от лишней массы, тем самым избавляется и от связанного с ней торможения роста, и период удвоения массы возвращается к своему первоначальному значению а0. Т:о., клеточный цикл, т.е. период времени от того момента, когда дочерняя клетка, образовавшаяся после митоза материнской клетки, начинает расти и до того момента, когда она сама став материнской клеткой, разделится на две дочерние, для популяции одноклеточных сохраняет постоянное значение.
В многоклеточном организме между образовавшимися после митоза клетками сохраняется связь, и по этой причине замедление роста возрастает от деления к делению.
Графически это показано на рис.1.
![]()
![]()
![]()

а) рост одноклеточных организмов
![]()
![]()
![]()
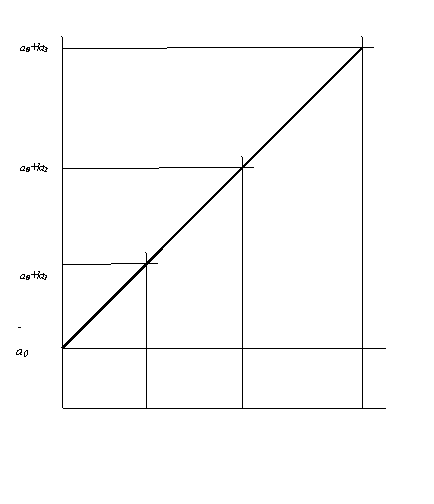
б) рост многоклеточного организма
Рис.1
Вышеизложенные рассуждения подводят нас к выводу о том, что замедление роста организма связано с нарастанием его массы. В пользу этого вывода свидетельствует следующий факт. Если сравнивать эмбрионы разных организмов одного и того же возраста, то относительный суточный прирост будет больше у того эмбриона, у которого масса на начало суток меньше. Это видно из приведенных таблиц. Все данные для таблиц взяты из книги акад. И. И. Шмальгаузена «Рост и дифференцировка».
Перепел Чибис
Возраст, сутки |
Масса, г |
Относительный прирост, % |
Возраст, сутки |
Масса, г |
Относительный прирост, % |
|
4 |
0,0196 |
4 |
0,0134 |
|||
6 |
0,1098 |
460 |
6 |
0,0865 |
545,5 |
|
7 |
0,2028 |
84,8 |
7 |
0,1684 |
94,7 |
|
8 |
0,3411 |
68,1 |
8 |
0,2956 |
75,5 |
|
9 |
0,5349 |
56,8 |
9 |
0,4813 |
62,8 |
|
11 |
1,1347 |
112 |
11 |
1,1960 |
148,5 |
Курица Мышь домашняя
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
|
8 |
1,210 |
8 |
0,00008 |
|||
9 |
1,630 |
34,7 |
9 |
0,00147 |
1737 |
|
10 |
2,846 |
74,6 |
10 |
0,0086 |
485 |
|
11 |
3,961 |
39,2 |
11 |
0,0329 |
282 |
|
12 |
4,650 |
17,4 |
12 |
0,0762 |
131,6 |
|
13 |
7,430 |
59,8 |
13 |
0,1298 |
70,34 |
|
14 |
9,84 |
32,4 |
14 |
0,2288 |
76,27 |
|
15 |
11,00 |
11,8 |
15 |
0,3651 |
59,57 |
|
16 |
13,65 |
24,1 |
16 |
0,5926 |
62,3 |
|
17 |
16,50 |
20,9 |
17 |
0,8467 |
42,88 |
|
18 |
20,70 |
25,5 |
18 |
1,1900 |
40,55 |
Курица Мышь домашняя Морская свинка
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
||
16 |
13,65 |
16 |
0,5926 |
16 |
0,0022 |
|||||
17 |
16,50 |
20,9 |
17 |
0,8467 |
42,9 |
17 |
0,0050 |
127,3 |
||
18 |
20,70 |
25,5 |
18 |
1,1900 |
40,6 |
18 |
0,,150 |
200 |
||
19 |
27,80 |
34,3 |
19 |
0,035 |
133,3 |
|||||
20 |
33,20 |
19,4 |
20 |
0,080 |
128,6 |
|||||
21 |
41,00 |
23,5 |
21 |
0,140 |
53,6 |
Крыса Мышь домашняя
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
Возраст, сутки |
Масса, г |
Относительный суточный прирост, % |
|
13 |
0,040 |
13 |
0,1298 |
|||
14 |
0,112 |
180 |
14 |
0,2288 |
76,3 |
|
15 |
0,168 |
50 |
15 |
0,3651 |
59,6 |
|
16 |
0,310 |
84,5 |
16 |
0,5926 |
62,3 |
|
17 |
0,548 |
76,8 |
17 |
0,8467 |
42,4 |
|
18 |
1,00 |
82,5 |
18 |
1,19 |
40,6 |
Рассматривая приведенные таблицы, мы видим, что указанное выше соотношение (из равновозрастных эмбрионов больший суточный прирост массы имеют те, у которых масса на начало суток меньше) в большинстве случаев выполняется. В пределах же одного организма указанное соотношение не выполняется ввиду колебательного характера процесса роста.
Можно привести еще один факт, свидетельствующий о том, что масса организма является фактором, тормозящим его рост.
На стадии дробления, как известно, увеличения массы зиготы не происходит, рост клеток отсутствует, и даже общая масса клеток несколько уменьшается. В прямом соответствии с этим продолжительность клеточного цикла на этой стадии постоянна, минимальна по длительности и даже отмечены случаи, когда эта продолжительность в ходе процесса дробления несколько уменьшается. И только когда начинается рост клеток, т.е. увеличивается масса эмбриона, продолжительность клеточного цикла также увеличивается. Приведенный вывод о постоянстве клеточного цикла на стадии дробления зиготы взят мною из книги Н. Н. Ротт «Клеточные циклы в раннем эмбриогенезе животных».
Итак,
из изложенных фактов следует, что замедление
роста живого организма связано с его массой.
Исследуем характер этой зависимости. Введем
величину, обратную периоду удвоения массы ![]()
Назовем ее потенциалом роста; она показывает как быстро происходит удвоение массы.
Построим график функции q=f(M)
Для
популяции одноклеточных
постоянная величина ![]() , не зависящая от массы M.
Для многоклеточных организмов зависимость q=f(M) имеет однотипный
характер, изображенный на рис.2б).
, не зависящая от массы M.
Для многоклеточных организмов зависимость q=f(M) имеет однотипный
характер, изображенный на рис.2б).
Примеры зависимости q=f(M) для отдельных многоклеточных организмов приведены на рис.3.

а) Рост популяции одноклеточных организмов
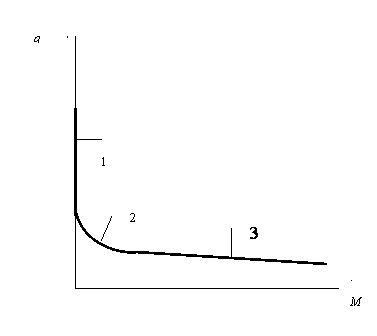
б) Рост многоклеточного организма
Рис. 2
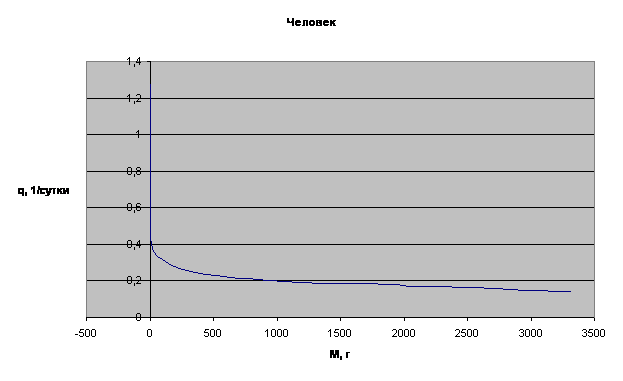
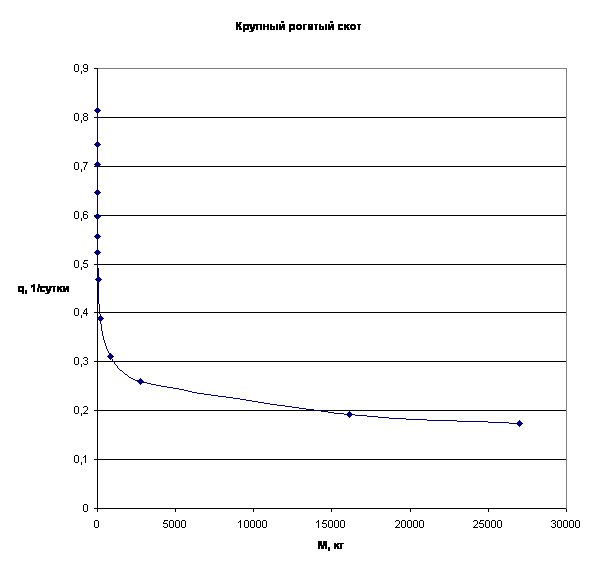
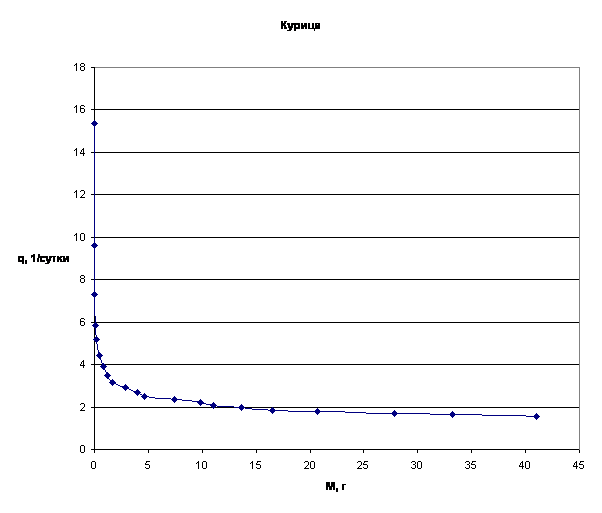
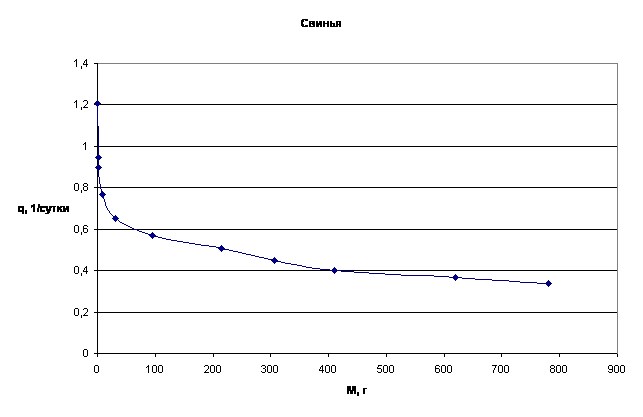
Рассматривая приведенные графики, мы приходим к выводу о том, что зависимость q=f(M) имеет характер короткодействия.
Представим себе некоторую среднестатистическую клетку организма окруженной определенной областью (для упрощения сферой определенного радиуса). Та масса, которая попадает в эту область, оказывает тормозящее действие на рост рассматриваемой клетки. Масса же клеток, находящаяся за пределами указанной сферы, никакого тормозящего действия на рост нашей клетки не оказывает. Т.е., фактор, тормозящий рост, резко обрывается за пределами определенной области, окружающей клетку.
График q=f(M) нам ничего не может сказать о природе этого фактора. Нельзя сказать, имеет ли он химическую природу (типа кейлонов), физическую природу (типа каких-либо излучений) или неизвестную нам природу (типа клеточных биологических полей А. Г. Гурвича). Ясно только, что этот фактор является короткодействующим по отношению к массе организма.
Аналогию короткодействия из физики мы усматриваем в поведении нуклонов в атомном ядре. Там поведение каждого нуклона определяется только теми нуклонами, которые находятся в непосредственной близости от него.
Короткодействующими являются также химические связи между молекулами твердых и жидких тел.
В свете вышеизложенного процесс роста многоклеточного организма можно представить следующим образом.
В результате процесса дробления зиготы образуется большое количество бластомеров. Каждый из них, являясь самостоятельной клеткой, имеет свою сферу короткодействия. Эти сферы ввиду тесного расположения перекрываются между собой. В результате, когда начинается рост бластомеров, каждый из них, испытывает на себе тормозящее действие со стороны всех остальных. Это приводит к резкому падению потенциала в начале роста (участок 1 на рис.2).
По мере роста организма все большее количество клеток переходит в такое состояние, когда сфера их короткодействия полностью заполнена массой других клеток. Назовем такое состояние клетки устойчивым или стационарным. Дальнейшее нарастание массы уже не может привести к снижению потенциала роста такой клетки. Этим объясняется явление насыщения. (участок 3 на рис.2)
Теперь поставим вопрос: что произойдет со стационарной клеткой, если часть массы из сферы ее короткодействия удалить? Как прореагирует на это клетка?
Клетка «почувствует» ослабление торможения роста и ответит на это делением клеток и их ростом. Это будет продолжаться до тех пор, пока не заполнится сфера короткодействия. Такое явление происходит при регенерации, как физиологической, так и репаративной.
Короткодействием фактора, тормозящего рост, можно объяснить, почему набольшая ранка, порез зарастает полностью, а например, отрезанный палец не отрастает.
Как в первом, так и во втором случае происходит нарастание массы в области короткодействия клеток раневой поверхности. Но в первом случае эта область полностью покрывает рану ввиду ее малого размера. Во втором случае нарастает только небольшой слой клеток вблизи поверхности раны, т.к. эта поверхность не воспринимает утрату клеток, находящихся за пределами области короткодействия.
Одноклеточные организмы, как не испытывающие торможения роста со стороны других клеток, обладают высокой регенерационной способностью.
Вопрос о том, что представляет собой область короткодействия, с помощью которой клетка воспринимает тормозящее рост действие со стороны других клеток, остается открытым. Может это и есть то клеточное биологическое поле, существование которого предполагал А.Г.Гурвич?
Продолжим наши рассуждения дальше и зададим вопрос: что произойдет, если по какой-либо причине область короткодействия какой-либо клетки окажется пустой?
«Почувствовав» свободу, такая клетка получает возможность бесконтрольного размножения. Если эта клетка внутренняя, то соседние клетки не дадут ей реализовать такую возможность. А если эта клетка поверхностная? Тогда ничто не мешает ее росту и размножению. Возникает злокачественная опухоль, которая растет по закону роста популяции одноклеточных, а сам организм для этой популяции уже является как бы питательным субстратом. С данным предположением хорошо согласуется тот факт, что злокачественный рост возникает только на покровных и соединительных тканях.
Т.о. с точки зрения предложенной гипотезы причина злокачественного роста состоит в нарушении нормальной интеграции поверхностных клеток, в результате которого отдельная клетка может освободится от тормозящего рост действия других клеток.
Такое нарушение может произойти под влиянием самых разных причин: механического удара, трения о различные предметы, воспалительных процессов, действия различных химических веществ, радиации и т.д.
Можно предположить также происшедшее под влиянием каких-то причин уменьшение радиуса короткодействия клетки, приведшее к тому, что масса других клеток, нормально тормозящая рост данной клетки, оказалась за пределами этого радиуса.
Может быть именно поэтому известно так много причин, вызывающих злокачественный рост. Причин много, а суть одна.
Доктор биологических наук А.Лучник сравнивает злокачественную опухоль с незаживающей раной. (Белорусская виртуальная библиотека. http://www.library.by/data/007/034.htm. Статья «Формула рака»).
Нормально заживление раны заканчивается как только заполнится область короткодействия клеток раневой поверхности, т.е. восстановится нарушенное равновесие. В случае злокачественного роста эта область не заполнится никогда, т.к. нарастание клеток хаотично и в пространственном отношении не подчинено никакой закономерности.
В заключение подведем итоги.
Доводы, приведенные в настоящей статье свидетельствуют о том, что замедление роста каким-то образом непосредственно связано с нарастающей массой организма. Но почему же тогда в формуле замедления периода удвоения массы, полученной в первой статье, масса отсутствует. По простой причине, по причине короткодействия фактора, тормозящего рост. Формула a=a0+kt характеризует рост организма на участке 3 рис.2, когда большинство клеток перешло в стационарное состояние, т.е. окружено тормозящей рост массой других клеток и от дальнейшего нарастания массы уже практически ничего не зависит (участок насыщения). Поэтому массы и нет в данной формуле. На этом участке все зависит от радиуса короткодействия и времени, что и отражает формула a=a0+kt . Величина k здесь не есть радиус короткодействия, но возможно с ним как-то связана.
Влияние нарастающей массы на рост многоклеточного организма, и при том очень сильное, имеет место только в самом начале эмбрионального роста, когда у большинства клеток области их короткодействия еще не заполнены тормозящей рост массой других клеток. (участок 1 на рис.2) По мере этого заполнения влияние нарастающей массы на замедление роста уменьшается (участок 2) и затем практически сходит на нет (участок 3).
Из-за сильного влияния массы на замедление роста на участке 1 зависимость a(t) отклоняется от прямой линии. Но этот участок очень непродолжителен по сравнению со всем периодом роста организма. Поэтому можно сказать, что исключая указанный участок, весь рост организма достаточно точно характеризуется формулой a=a0+kt . Каков ее смысл? Она выражает возрастное старение организма. Все изложенное подводит к следующей гипотезе. С каждой единицей живой материи в каждую единицу времени связано что-то, что замедляет процессы биосинтеза в организме, причем как на уровне одной клетки, так и на уровне многоклеточного организма. Видимо, живое не может существовать не старея и старение есть такое же свойство живой материи, как и все другие известные ее свойства.
Если принять это положение, то как следствие остается признать, что никакого бессмертия на уровне отдельного организма быть не может. Можно только говорить об относительном «бессмертии» популяции, «бессмертии» пока для существования этой популяции есть необходимые условия.
В применении к человеку этот вывод согласуется с выводом геронтологов о том, что по мере улучшения социальных условий максимальный предел жизни человека не увеличивается, а только увеличивается количество людей, приближающихся к этому пределу.
Литература
1. Ротт Н.Н. Клеточные циклы в раннем эмбриогенезе животных. – М.:Наука, 1987. – 287 с.
2. Свечин К.Б. и др. Возрастная физиология животных – М: Колос, 1967 – 430 с.
3. Шмальгаузен И.И. Рост и дифференцировка. Т.1. – Киев: Наукова думка, 1984. – 176 с.
4. Шмальгаузен И.И. Рост и дифференцировка. Т.2. – Киев: Наукова думка, 1984. – 168 с.
The analysis of the
cause of delay of living organisms’ growth
UDK 577/95
Sedova G.P.
This
article contains an attempt to explain the cause of delay of multicellular
organisms’ growth.
The analysis
of universally known facts draws a conclusion that a factor which
causes the delay of growth is somehow, connected with a going mass of organism.
Graphic
representation of this dependence dives evidence of short action of this factor relating
to a mass. It means that every cell of organism receives a delaying action only from the
side of neighbouring cells.
The phenomenon of
regeneration is also can be explained by a short action of growth delaying factor. In
wound place only the coat of cells grows which is situated in
radius of short action of wound surface cells.
Wound surface cells
do not receive the loss of cells situated beyond the radius of short action so that is why
these cells are not regenerated.
A hypothesis of
malignant growth can be suggested, taking as the basic the short action of the growth delaying factor.