УДК: 572.
018.722
ББК: 28.71
Симметрия и сегментация в структурной организации конечностей
человека
Ó
2006 г. Ермоленко А. Е., Перепада Е. А.
Предложена оригинальная модель развития
конечностей человека. В основе развития конечностей человека лежит метамерная
закладка костных отростков в соответствии с первичной сегментацией и
двухплоскостной симметрией. Костные отростки изначально идентичные друг другу в
процессе эволюции не пропорционально изменялись в размере,
перегруппировывались, объединялись между собой, регрессировали сохраняя направленность построения по схеме:
в верхней конечности вместе с плечевым поясом (с учетом регрессировавших
и сесамовидных костей) - 2; 1; 2; 3; 2; 3; 5; 5; 8; 8; в нижней конечности
вместе с костями таза (с учетом регрессировавших и сесамовидных костей) - 3; 2; 3; 2; 1; 2; 8; 8; 5; 5.
Введение
Определение закономерностей структурной
организации в эволюции человека как биологического вида являются одной из
важнейших проблем биологии. Существуют различные точки зрения в изучении
механизмов развития конечностей. Если однако, факт
происхождения этих конечностей от плавников является несомненным, то остается
все же еще вопрос - каков же тип того плавника, из которого развилась пятипалая
конечность? В общем, все решения вопроса группируются около двух точек зрения.
Одни считают, что конечность наземного позвоночного произошла от плавника,
имевшего лишь один луч внутреннего скелета, другие, что конечность наземных
позвоночных произошла из типичного рыбьего плавника, обладавшим не особенно
большим числом лучей [10].
Автором данной работы
предложена собственная концепция развития конечностей человека. Но для рассмотрения
этой концепции необходимо сделать отступление и принять к сведению гипотезу о
биокристаллоидном устройстве биологических организмов [14], которое
обеспечивает симметрию и сегментацию организма, обратить внимание на некоторые
этапы эволюционного развития далеких предшественников человека.
Теортические предпосылки
Организм человека является
биокристаллоидом, который следует понимать в виде композиции кристаллоподобной
структуры и околокристаллической среды, как на уровне целостного организма, так
и на уровне клеток. Анализ биологических и небиологических минералов показывает, что они кристаллографически не
имеют фундаментальных различий, что говорит о функционировании общих
кристаллизационных процессов [1, 3, 12,
13]. Главной характеристикой кристаллических структур является их симметрия.
Для человека свойственна двухплоскостная симметрия, которая обусловлена
двухплоскостной симметрией хромосомы – носительницы генетической информации.
Одной из фундаментальных особенностей минерального вещества является его
агрегация, т.е. существование минеральных
индивидов не только отдельно друг от друга, но в виде агрегатов, закономерных срастаний
индивидов, а также других синминералогических систем. Это свойство присуще не только сложным молекулам,
но и ассоциатам более высокого порядка. Из биологии клетки [8] известно,
что при соприкосновении однотипных клеток друг с другом они слипаются, образуя агрегаты
характерные для данного вида клеточных популяций. Способность к агрегации не
хаотично, а в определенной последовательности выражается в самоорганизации. Самоорганизация живой материи, формообразование тканей в
значительной мере зависит от межклеточной самосборки, в которой участвуют как
простые молекулы, так и макромолекулы. Процесс объединения клеток
контролируется молекулярными посредниками и реализуются при участии
межмолекулярных взаимодействий [6].
Интеграция
- способность живых однотипных систем и их частей, совместно выполняющих
одинаковую функцию, объединяться сначала в конгломерат (колонию), а затем в
единый организм. Это могло происходить при образовании из отдельных клеток
шаровидного организма типа «volwox» - сначала
колония, а затем единый организм, это же явление характерно и для
многоклеточных организмов. Интеграцией объясняется и объединение в единый
организм многосегментных особей, с последующим созданием организмов, состоящих
из двух групп сегментов, т.е. создание организма из двух разных, но однотипных
организмов, каждый из которых имел разное количество сегментов. Сегментация -
это не что иное, как следы границ после интеграции в единый организм отдельных
многоклеточных несегментированных особей с постепенным сглаживанием между ними
разницы и создание единого организма по законам формообразования. Кроме того, в
результате изменения количества плоскостей симметрии при переходе от
трехплоскостной к двухплоскостной симметрии не все организмы одинаково
построили антимеры. Одна группа животных построила их в ряд, соблюдая
метамерию, при этом метамерный ряд увеличился (к этой группе относятся, в
частности, хордовые), а другая объединила антимеры, соблюдая первичную
сегментацию (к этой группе относятся членистоногие).
Нами [15] предложена гипотеза, объясняющая
образование сегментированных животных путем агрегации простых не
сегментированных биокристаллоидных организмов, сначала в колонию, а в
дальнейшем в единый сегментированный организм, с медленным стиранием граней
первичной агрегации. В филогенезе далекий
предшественник человека прошел развитие от простой клетки через этапы
клеточного колониального сообщества, многоклеточного, несегментированного
организма типа «вольвокс»; организма типа "вогнутого мяча", на
подобии гастреи Э. Геккеля или паренхимеллы И. Мечникова, колоний
многоклеточных несегментированных организмов до образования цельного
многосегментного организма. Вероятно, наиболее устойчивой формой колоний было
небольшое количество организмов, что привело к созданию от 2 до 9 сегментных
организмов. Из этих многосегментных организмов более распространенной формой
были 5 сегментные. В дальнейшем шло объединение не только отдельных особей, но
и групп. Как сообщает Я. И. Старобогатов [9], например, членистоногие состоят
из двух групп сегментов, при этом головная группа всегда состоит из 5, а
каудальная от 2 до 9 сегментов. Интеграция 5 и 8 сегментных организмов привела
к образованию 13 сегментного организма-предшественника человека.
Сегментация
конечностей
Современные биология и анатомия не дают
четких представлений о сегментации животных и о ee происхождении. Человек считается частично сегментированным.
Нет четких представлений о количестве сегментов, присущих человеку. Нет
согласия среди ученых о количестве сомитов у человека – их насчитывают от 37 до
44 [2]. П. П. Иванов [4] приводит сравнительный анализ метамерии в
эмбриональном развитии разных животных. Однако он не предложил никаких идей,
объясняющих метамерию. Далее автор делает вывод, что хордовые могли произойти из
олигомерных форм. О причинах, заставляющих олигомерные формы превращаться в
полимерные, нет сведений. Не решен вопрос о происхождении наземных позвоночных
животных, имеется разногласие в вопросе о их монофилии или полифилии. N. Holmgren [16]
считает, что они имеют полифилитическое происхождение. А И. И. Шмальгаузен [11]
убежден в их монофилитическом происхождении. При этом одним фактам и доводам
придается более важное значение, а важные доводы оппонентов занижаются. Однако
вопрос надо поставить шире. Объясняется ли разнообразие строение тела животных только
дивергенцией, обусловленной условиями существования? На наш взгляд вопрос может
быть решен на основании анализа сегментного строения целостного организма. С
решением этого вопроса тесно связана проблема образования новых органов. Р.
Кэрролл [5] говорит: «Нужно предполагать, что метакарпалии и фаланги пальцев
тетрапод развились, если не совсем, то почти полностью как новообразование».
Однако новые органы не образуются из ничего (ad novo). Они
могут образовываться из уже имеющихся органов или их частей с приобретением
новых функций и последующим обособлением. Разделение имеющихся органов
происходит в соответствии с присущей данному типу организмами симметрией и
сегментацией (для позвоночных – это четырехсторонняя симметрия). При этом у
вида, находящегося в начале эволюционного процесса, органы (в частности кости)
могут находится в зачаточном состоянии или вовсе не проявляться, а у вида,
находящегося в конце его, части органов интегрируются, редуцируются и исчезают.
Скелет парных плавников возник в виде
метамерного ряда хрящевых стержней в первоначально непрерывных парных боковых
складках. Путем разделения этой складки затем образовались отдельные пластинки,
причем на каждый метамер туловища приходится два луча парного плавника [10].
Путем перегруппировки закладок с развитием конечных частей складок и редукции
промежуточной области образовались передние (верхние) и задние (нижние)
конечности. Метамерная закладка мускулатуры и скелета служит указанием на то,
что этот непрерывный плавник был построен метамерно, в точном соответствии с
сегментацией туловища [10]. Разделению общего плавника на отделы соответствовало
несомненно и физиологическое разделение функций.
Из непрерывной боковой складки
образовалось 4 группы костей в соответствии с двухплоскостной симметрией и 13
сегментным делением. В каждой группе было по 52 закладки, которые служили в дальнейшем
материалом для образования парных плавников, а затем и конечностей. Редукция
количества закладок в конечности с 52 до 39 связана, вероятно, с интеграцией и
деградацией элементов антимеров. Из костных закладок каждой группы
образовалось два отдела: проксимальный и дистальный. Первый имел один ряд (13
закладок), состоящий из духа групп (5 и 8 закладок), второй из 2 двухгруппных
рядов (по 13 закладок) (рис. 1). Последующая трансформация привела к
образованию удлиненного органа движения, у которого было 6 рядов (два ряда проксимального отдела и 4
ряда дистального) по схеме: 5; 8; 5; 5; 8; 8 (для передней конечности) или 8;
5; 8; 8; 5; 5 (для задней конечности). Наряду с увеличением количества рядов в
конечности кости, образовавшиеся из изначально равнозначных закладок, приобрели
различные размеры. Дальнейшее усложнение привело к распределению костей
проксимального отдела в 6 компонентной последовательности, дистальный отдел
остался состоять из четырех рядов. При этом кости проксимального участка верхней
конечности формировались по схеме: 2; 1; 2; 3; 2; 3, а нижней конечности по
схеме: 3; 2; 3; 2; 1; 2. Часть костей конечности из-за морфогенетических
воздействий преобразовались в сесамовидные кости или редуцировались полностью,
что изображено на нашей схеме (рис.1) укороченными штрихами. Иногда
сесамовидные кости получают дополнительное развитие, перегруппировываются,
образуя, как бы семилучевой веер на руке, однако общее количество костей,
включая сесамовидные, не превышает количество первичных закладок, которые
соответствуют количеству первичных сегментов при двухплоскостной симметрии.
Можно
предположить, что реальное животное, у которого конечности были бы построены по
описанной схеме, никогда не существовало, так как непрерывно шла их
реорганизация. Эти схемы предложены для оценки направленности процессов.
Костные закладки из непрерывной боковой складки многократно
перегруппировывались, организовывались в восьмипалую (переднюю) и пятипалую
(заднюю) конечность. Дальнейшие перегруппировки были связаны со стремлением
организма построить идентичные антимиры (пятипалые конечности). Предложенные
схемы показывают разную судьбу закладок 5 и 8 сегментных организмов,
послуживших материалом при формировании конечностей. Реальное строение руки
человека (вместе с плечевым поясом) практически полностью совпадают с
предложенной схемой. Реальное строение ноги человека (вместе с костями таза)
можно считать соответствующим схеме при определенных допусках. Такая трактовка
распределения костей в организме по данной схеме, вероятно, будет принята не
сразу, однако если учесть ряд факторов, можно предположить, что предложенная
схема верна. По А. Ромеру и Т. Парсонсу [7] пяточная кость (os calcaneus) филогенетически относятся к голени. На рис. 2 представлена
схема расположения берцовых и пяточной костей у млекопитающих по А. Ромеру и Т.
Парсонсу. У предков млекопитающих кроме
типичных трех костей таза развивается еще одна, [5, 7, 10]. Вероятно, центр
формообразования этой кости сместился и образовал вторую кость бедра, которая
затем сместилась ниже и стала таранной костью (os talus). В
процессе передвижения этой кости была смещена ниже третья кость голени
(пяточная) и обе кости стали составной частью стопы.
На
рис. 3 представлена схема строения конечностей акантостеги (acanthastega gunnari), по J. Clack [17, 18], созданной на основании ископаемых
окаменелостей. Восьмипалая передняя
конечность acanthastega
gunnari вполне подходит под
предложенную нами схему (рис.1). Надо обратить внимание, что строение передней
и задней конечностей acanthastega
gunnari различно, что также соответствует
нашей схеме.
Заключение
При формировании верхней конечности
человека в ее состав вошли костные закладки проксимального участка и образовали
13 костей: лопатку (os scapula), ключицу (os clavicula), плечевую (os humerus),
лучевую (os radius), локтевую (os ulna) и все 8 костей запястья (ossa carpi).
Также шло формирование нижней конечности и были образованы 13 костей:
подвздошная (os ilium), лобковая (os pubis), седалищная (os ischii), бедренная
(os femoris), малая (os fibia) и большая (os tibia) берцовые и 7 костей
предплюсны (ossa tarsi). Дистальные закладки образовали на руке кости пясти
(ossa metacarpi), фаланги пальцев (phalanges); на ноге - кости плюсны (ossa
metatarsi) и фаланги пальцев (phalanges). Остальные закладки не участвующие в
органе движения, получили обратное развитие - регрессировали или представлены в
виде сесамовидных костей. Максимальное количество сесамовидных костей 28 (по 7
на каждую конечность с учетом неразвившихся фаланг первых пальцев). Надо
отметить, что расположение костей в запястье и в предплюсне не позволяет
достоверно определить их рядность.
Изучение строения конечностей человека
позволяет сделать вывод о чередовании групп, состоящих из 5 и 8 костных
закладок. В верхней конечности проксимально включена группа из 5 костей
(лопатка, ключица, плечо, лучевая, локтевая кости) затем группа из 8 костных
закладок (кости запястья), затем снова два ряда по 5 костных закладок (кости
пясти и один проксимальный ряд фаланг пальцев). Заканчивается верхняя
конечность двумя рядами по 8 костных закладок, у которых в каждом ряду по 3
кости регрессированы. На нижней конечности
проксимально включена группа из 8 костных закладок (в противоположность верхней
конечности, где проксимально включены кости пятисегментного организма) это три
кости таза, бедренная, берцовые кости, таранная и пяточная. Далее расположена
группа из пяти закладок (ладьевидная –os
naviculare,
кубовидная – os cuboideum и три клиновидных ossa cuneiformia) и снова два ряда по 8 костных закладок. Заканчивается
нижняя конечность двумя рядами по 5 костей. На первом пальце всех конечностей
средняя фаланга регрессировала.
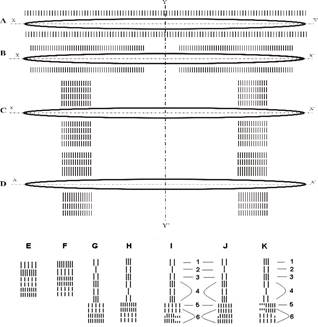
Рис. 1.
Схема образования конечностей человека
x - x’; y – y’ – плоскости симметрии.
A - парная непрерывная боковая складка.
B - D - поэтапное образование антимеров с костными закладками в двухплоскостной симметрии, служащими материалом для формирования конечностей.
B - образование антимеров с расположением закладок в ряд.
C - перегруппировка закладок.
D - частичная интеграция закладок в антимере.
E - K - последовательное формирование конечностей и их поясов.
E, G, I, J - образование верхней конечности; J - семилучевой орган движения.
I, J: 1 - кости плечевого пояса; 2 - плечо; 3 - предплечье; 4 - запястье; 5 - пясть; 6 - фаланги пальцев руки.
F, H, K - образование нижней конечности.
K: 1 - кости таза; 2 -
бедро и таранная кость; 3 - берцовые и пяточная кость;
4 - предплюсна; 5 - плюсна; 6 - фаланги пальцев ноги.
I, J, K - укороченные штрихи обозначают редуцированные и сесамовидные кости.
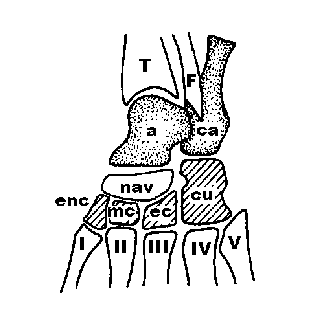
Рис.
2. Схема предплюсны древних млекопитающих по А. Ромеру и Т. Парсонсу [7].
T –
tibia; F – fibula; ca – calcaneus; a – astragalus; nav – naviculare;
mc – mesocuneiforme; ec - ectocuneiforme; cu -cuboideum.
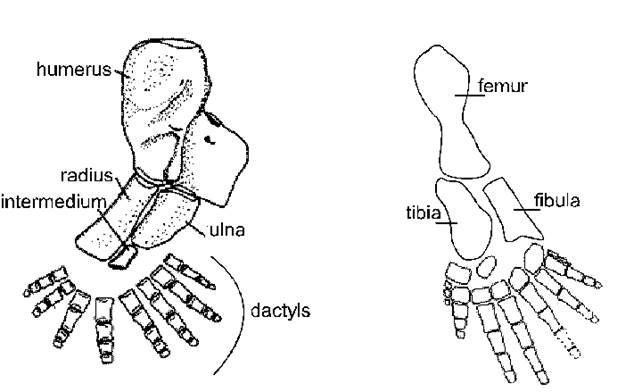
Рис. 3. Левые конечности
Acantostega по J.Clack [17].
Литература
1. Асхабов А. М. Кваторонная природа морфологической схожести биогенных и абиогенных наностуктур. //Минералогия и жизнь: биоминеральные гомологии. - Республика Коми, Сыктывкар: Геопринт, 2000. - С.12-14.
2.
Борзяк Э.И. с
соавт. Анатомия человека. – М.:
Медицина, 1986.
3. Войтеховский Ю. Л. Биоминеральные гомологии. //Минералогия и жизнь: биоминеральные гомологии. - Республика Коми, Сыктывкар: Геопринт, 2000. – C. 18-22.
4.
Иванов П. П. Общая и сравнительная эмбриология. -
Москва- Ленинград: ОГИЗ – БИОМЕДГИЗ, 1937. - C. 730-733.
5. Керролл Р. Палеонтология
и эволюция позвоночных. (Carroll R.
Vertebrate Paleontology and Evolution). – М.: Мир, 1993.
6. Лима-де-Фариа
А. Эволюция без отбора. Автоэволюция формы ифункции. (Lima-de-Faria
A.Evolution without Selection. Form and Function by
Autoevolution. Elsilver. Amsterdam, New York. Oxford). – М.: Мир, 1991.
7. Ромер А.,
Парсонс Т. Анатомия позвоночных. (Romer A.,
Parsons T. The vertebrate body.). – М.: Мир, 1992. –
C. 254.
8. Свенсон К.,
Уэстер П. Клетка. (Swanson C. P., Webster R. L.
The Cell). - М.: Мир, 1980.
9. Старобогатов Я. И. Филогения и система членистоногих (ARTHROPODA). Успехи современной биологии. - Том III. – Вып. 6. - 1991. - С. 828-839.
10.
Шмальгаузен И. И.
Основы сравнительной анатомии позвоночных животных. М.: Советская наука,
1947.
11.
Шмальгаузен И. И. Происхождение наземных позвоночных.
– М.: Наука, 1964.
12.
Юшкин Н. П. Биоминеральные взаимодействия. – М.:
Наука, 2002.
13. Юшкин Н П. Биоминеральные гомологии и организмобиоз. //Минералогия и жизнь: биоминеральные гомологии. – Сыктывкар: Геопринт, 2000. - С. 9-12.
14. Ermolenko A.E. Two-plane symmetry in the structural
organization of man. Medical Hypotheses (2005) 64, 209-214.
15. Ermolenko A., Perepada E. Origin of
segmentation in the human structure. Medical Hypotheses. (2006) 67,
622-625.
16. Holmgren N. Contributions to the
question of the origin of the tetrapods. Acta Zool., (1949) 30, №
3, 89 – 124
17. Clack, J. A. An
Early Tetrapod from "Romer's Gap." Nature (4 July, 2002) 418,
72-76
18. Clack, J.A., Gaining Ground: The Origin and Evolution
of Tetrapods, Indiana University Press, Bloomington, 2002. www.answersingenesis.org/tj/v17/i2/tetrapod.asp
Symmetry and segmentation in the
human limb structure
Ermolenko
A. E., Perepada
E. A.
We propose an
original model of human limb evolution. The human organism is a biocrystalloid
with a structure comparable to that of mineral organisms with symmetry and
segmentation typical of them. One of the fundamental characteristics of
individual mineral specimens and biocrystalloids is their ability to aggregate,
i.e. to exist not only as separate objects but also in
the form of aggregates and regular concretions. Segmentation is nothing else
but traces of boundaries left following integration of separate multicellular
non-segmented specimens with gradual fading of their differences and formation
of a new entity in compliance with formation laws. Integration of five- and
eight-segment organisms resulted in the formation of a 13-segment precursor
organism of man. Human extremities developed from the symmetrical metameric
anlages of bone processes in conformity with the original segmentation and
two-plane symmetry. In the process of evolution originally identical limb bone
processes disproportionately changed in size, regrouped, united, regressed,
resulting in arrangement according to the schemes: for the shoulder girdle and
the superior extremity - 2; 1; 2; 3; 2; 3; 5; 5; 8; 8; and 3; 2; 3; 2; 1; 2; 8;
8; 5; 5 for the pelvis bones and the inferior extremity. This includes reduced
and sesamoid bones as well.
Key words: tetrapods, limb development, symmetry,
segmentation.
ФГУ НИИ
трансплантологии и искусственных органов Росздрава
Поступила в
редакцию 27.10.2006.