УДК 615.015
МОДЕЛИРОВАНИЕ ФИЗИКИ МОЗГА
© 2006 г. Холманский А. С.
Физика мозга человека имеет две составляющие – базовую физику общую для всех млекопитающих и физику мышления, присущую только человеку. Развитие ментальной составляющей структурно-функциональной организации мозга в филогенезе связали с хиральным фактором внешней среды, а в онтогенезе – с социальным фактором. В основу чувствительности мозга к данным факторам положили односвязность его водной основы, механизм электромагнитной индукции и особенности термодинамики мозга в состоянии ночного сна. С целью унификации описания механизма электромагнитных процессов в мозгу ввели понятие квазифотона, объединяющее в себе все формы возбуждения электронных и молекулярно-клеточных структур мозга. Предложены эквивалентные схемы колебательных контуров элементов нейросети и макроструктур мозга. Сделаны оценки кинетических параметров (энергии активации, скорости) физических процессов, лежащих в основе энергоинформационного обмена мозга с внешней средой. Обсуждены механизмы работы оперативной (физической) и постоянной (химической) памяти мозга, включая модель нелокальных квантовых корреляций.
Оглавление
1. Концептуальное введение
2. Электромагнитная индукция
2. 1. Колебательный контур
2. 2. Электромагнетизм нейрона
2. 3. Нейросети
2. 4. Капсулированные нервные окончания
2.4.1. Тельце Фатера Пачини
2.4.2. Глаз
3. Квазифотон
3.1. Типы квазифотонов
3.2. Метрика квазифотона
3.3. Метаболические квазифотоны
4. Термодинамика мозга
5. Физика организации мозга
5.1. Функциональная иерархия мозга
5.1.1. Кора больших полушарий
5.1.2. Таламус, желудочки мозга
5.1.3. Эпифиз
5.2. Кинетические параметры физики мозга
5.3. Нелокальные квантовые корреляции
5.3.1. Сознание и память
5.3.2. Механизм квантовых корреляций
6. Заключение
Литература
1. КОНЦЕПТУАЛЬНОЕ ВВЕДЕНИЕ
Основным структурно-функциональным элементом мозга является нервная клетка. Она генерирует и проводит электрические импульсы – потенциалы действия (ПД). Связанное с ПД движение зарядов индуцирует локальные вихри электромагнитного (ЭМ) поля, которые, в принципе, можно определить как ЭМ-кванты или квазифотоны. Метрика, принцип и скорость движения квазифотона будут определяться электрофизическими свойствами и структурными особенностями нейрона и окружающей его среды. За энергоинформационное обеспечение механизма генерации импульсов и за синтез метаболитов ответственно тело клетки, ее ядро и дендриты. Аксоны в симбиозе с нейроглиями (олигодендроциты, астроциты) транслируют метаболиты и импульсы, реализуя их энергию и информацию через синтез и действия нейромедиаторов в синапсах. Дееспособность нервной клетки обеспечивает энергия реакции окисления глюкозы, которая в митохондриях трансформируется в энергию макроэргических связей АТФ. В нервных клетках энергия АТФ преобразуется в энергию квазифотонов, в энергию химических связей синтезируемых веществ, в кинетическую энергию метаболитов и молекул среды (тепло). За счет этой же энергии осуществляется рост аксонов, развитие нейронных сетей и нейроглиальных связей, которые, в частности, отвечают за механическую целостность цитоскелета мозга. Физико-химические свойства воды, составляющей основу жидкостных систем мозга (ликвора, крови), в полной мере ответственны за электрофизику мозга и за его термодинамические свойства, как на микро, так и на макро уровнях его организации.
Таким образом, поведение мозга как единой физической системы в первую очередь подчинено классическим законам электрофизики и термодинамики сплошных коллоидных сред. В рамках данных законов осуществляется метаболизм нейрона, и мозг исполняет свои базовые функции, управляя гомеостазом и своевременно запуская механизм полового размножения. Соответствующая данным функциям физика мозга будет одинакова для всех млекопитающих, поэтому ее можно считать базовой. Именно это и позволяет экстраполировать результаты исследования мозга животных на мозг человека. Однако только организм гоминида (homo erectus) на этапе прямохождения приобрел чувствительность к фактору филогенеза геокосмического масштаба [1], под влиянием которого в условиях социальной среды в его мозгу стали формироваться и развиваться структуры ответственные за речь и мышление. Анатомические различия мозга современного человека и обезьяны ярко выражены в строении и объеме лобно-височных долей неокортекса. Ключевую роль в физике мышления играет структурно-функциональная асимметрия полушарий мозга, которая отсутствует у животных и имеет расово-половую дифференциацию у человека. Генезис данной асимметрии мог быть детерминирован перестройкой физики половых органов, рук, зрения и слуха на этапе прямохождения и в процессе развития навыков к сознательному труду. Исходя из этих данных, в основу когнитивных функций мозга положим физику лобно-височных долей неокортекса и хиральность коммуникаций мозга, как межполушарных и соматических, так и с внешней средой.
Учитывая наличие в мозгу метастабильных и динамичных квазифотонов различных типов и энергий, можно предполагать их активное участие не только в метаболизме, но и в физике когнитивных функций в рамках законов классической квантовой механики. Природа внешнего универсального хирального фактора, как и природа хиральных квантов энергии в мозгу не обязательно должна совпадать с природой квазифотонов, метрика которых, тем не менее, может быть спиральной. Механизмы поглощения и действия в мозгу хиральных квантов энергии (например, нейтринной природы [1]) тесно связаны с физикой самоорганизации и фазовых переходов в кооперативных хиральных системах [2, 3].
Главный вопрос физики мозга состоит в моделировании механизма психофизического изоморфизма [4], который, по сути, суммирует в себе следующие процессы:
– формирование на уровне атомно-молекулярной системы ЭМ-матрицы смысла-слова (мыслеформы);
– распознавание и вербализация другой системой атомов содержания мыслеформы.
Пространственно-временная разделенность двух физических систем, участвующих в формировании и распознавании мыслеформы предполагает физическое обособление мыслеформы в виде связанной системы дискретных форм материи, изоморфной ЭМ-матрице мыслеформы. Физическая обособленность мыслеформы является необходимым условием и для адекватности обмена информацией по механизму нелокальных квантовых корреляций. Идеальным, в этом смысле, носителем мыслеформы могут быть простейшие формы материи, предшествующие квантам полей и элементарным частицам. Тогда задача согласования и стыковки физики мышления с физикой базовых функций мозга сведется к проблеме вербализации фундаментальной динамической формы материи, способной благодаря своему движению становиться носителем энергии и информации [5]. Аксиоматику простейших форм материи (энергоформ) построили [6], опираясь на законы диалектики и экстраполируя достоверные положения классической и квантовой физики. Универсализм энергоформ (ЭФ) позволяет их использовать для моделирования мыслеформ, квазифотонов и предшественников элементарных частиц. Взаимодействия ЭФ с веществом мозга идут при посредничестве квазифотонов, сочетая фрактально-резонансный принцип действия ЭФ [6] с механизмом нелокальных квантовых корреляций [7].
К энергоформам и их конденсатам, по сути, относятся гипотетические “струны”, “кварки”, “вихри Абрикосова”, “матрицы плотности” и другие абстрактные модели субэлементарных дискретных форм материи. В работе [8], при анализе термодинамики мыслительной деятельности мозга, на роль “рабочего тела” аппарата мышления был предложен газ гипотетических х-частиц (фермионов), распределенный, по нейронной сети коры мозга. Если попытка отнесения х-частиц к нейтрино безосновательна [4, 6], то некоторые особенности термодинамики х-частиц приемлемы для биоактивных ЭФ и квазифотонов.
Таким образом, физику мышления можно обособить в рамках физики базовых функций мозга, отнеся к ее ведению уникальную способность вещества мозга при нормальных условиях резонансно поглощать, генерировать, селектировать, комбинировать и сохранять дискретные формы материи (энергоформы и квазифотоны), распознавая в их действиях смысл-слова, психическую или иную ментальную информацию. С целью обоснования применения энергоформ и квазифотонов для моделирования физики мышления в настоящей работе проанализировали структурно-функциональные особенности мозга и сделали оценки энергий активации (ЭМ-квантов) ключевых физико-химических процессов, обеспечивающих энергоинформационный обмен внутри мозга и между мозгом и внешней средой, к которой относится также и тело человека. Результаты анализа и оценок использовали для проведения экстраполяций известных физических закономерностей на уровень физики энергоформ.
2. ЭЛЕКТРОМАГНИТНАЯ ИНДУКЦИЯ
2. 1. Колебательный контур
Кинематику ЭФ [6] иллюстрирует явление электромагнитной индукции (ЭМИ), которое формально подчиняется первому уравнению Максвелла [9]:
rotE = - дB/дt , (1)
где Е и В – взаимно ортогональные вектора напряженности вихревых электрического и магнитного полей.
С помощью (1) для замкнутого контура с током получают уравнение для ЭДС самоиндукции (U):
U = – L (dJ/dt) = – dФ/dt, (2)
где L – индуктивность контура; J – ток, а Ф = LJ – потокосцепление самоиндукции контура.
Эффекты ЭМИ в различных структурах и средах живого организма, имеющих свои локальные магнитные (m) и диэлектрические (e) характеристики, подчиняются второму уравнению Максвелла:
rotH = j + дD/дt , (3)
где
D = eoe E , B = momH, (4)
j – ток смещения, а электродинамическая постоянная вакуума (eomo) и среды связаны со скоростями распространения ЭМ-квантов в вакууме (C) и среде (V) соотношениями [9]:
C = (eomo)–1/2 , V = С(em)–1/2 = С/n (5)
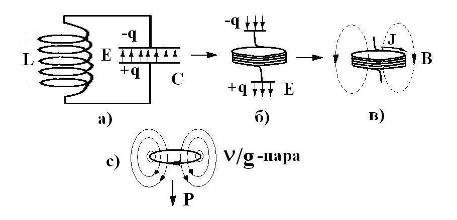
Экстраполяцию явления ЭМИ на уровень ЭФ [6] можно проиллюстрировать на примере колебательного контура (Рис 1).
Рис. 1. Колебательный контур – а) и его трансформированные формы, отвечающие началу колебаний – б) и четверти периода – в); с) – экстраполяция состояния контура в) на уровень энергоформы (n/g-пара), имеющей импульс Р и эквивалентную массу mg .
Для идеального контура частота гармонических электромагнитных колебаний задается формулой:
w = (LC)–1/2 (6)
Трансформация колебательного контура путем раскрытия конденсатора и сжатия катушки показана на Рис. 1. Состояние б) отвечает схеме антенны, которая может, в принципе, принимать и излучать фотоны радиоволнового диапазона. При этом вихревые Е и В-поля заполняют все пространство. Трансформация в) отвечает состоянию колебательного контура, когда энергия Е-поля перешла в энергию вихревого В-поля. Конфигурацию ЭМ поля в состоянии в) можно отождествить с ЭФ (n/g-пара [6]), связав ее импульс Р или энергию Е-поля, с импульсом тока до его закручивания в спирали катушки. Соответственно, вращательный момент тока или связанная с ним энергия В-поля будут отвечать моменту импульса ЭФ или ее эквивалентной массе (mg). При комбинации различных n/g-пар собираются кванты полей (фотоны, гравитоны), а при их конденсации числом, равным числу Авогадро (6 ·1023), образуются элементарные частицы [6].
2. 2. Электромагнетизм нейрона
Очевидно, что ЭМИ играет существенную роль в механизмах генерации и действия ЭФ электромагнитной природы в нервной системе человека. В основе ее коммуникативных и сигнальных функций лежит способность нервных клеток генерировать и проводить электрические импульсы. Электрофизику и метаболизм нервной системы и нейронов исследуют с помощью методов ЭКГ, ЭЭГ, МЭГ, ЯМР и позитронно-эмиссионной томографии (ПЭТ), термоэнцефалоскопии, психофармакологии и непосредственным зондированием нервных клеток микроэлектродами. Квантовые магнитометры (СКВИД), в принципе, позволяют регистрировать магнитное поле отдельного нейрона [10, 11]. Явления ЭМИ и резонанса, по-видимому, лежат в основе механизма чувствительности нервной системы к прямым воздействиям внешних ЭМ-излучений различного диапазона. Наличие в нервной системе LC-структур, в принципе, допускает “настройку” чувствительных элементов нервной системы на частоты как внутренних, так и внешних биогенных излучений по принципу гетеродинной связи.
Для объяснения электрических свойств мембраны привлекают схему эквивалентного контура, в которой проводящие каналы для различных ионов моделируют источником ЭДС и омическим сопротивлением (R), а изоляционные свойства мембраны представляют емкостью (Рис. 2).
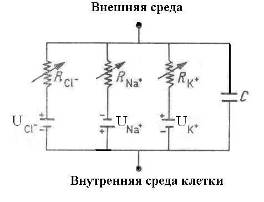
Рис. 2. Эквивалентная электрическая модель мембраны нерва: батареи создают суммарный мембраны потенциал U, ионная проводимость обозначена сопротивлениями R, емкость мембраны – конденсатор С [12].
Параллельное соединение нескольких контуров, показанных на Рис. 2, моделирует мембрану нейрона [12]. Однако для модели нейрона центральной нервной системы, имеющего миелиновую оболочку, емкостной характеристики мембраны не достаточно. Действительно, в спиральной структуре миелина есть регулярные каналы (насечки) (Рис. 3), которые в контексте эквивалентной электрической модели мембраны (Рис. 2) вполне могут играть роль локальных катушек индуктивности. Число насечек на одном миелиновом сегменте волокна, тем больше, чем толще осевой цилиндр аксона [13].
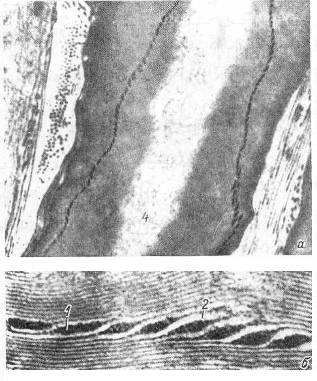
Рис. 3. Ультраструктура миелиновой мембраны нерва. а – общий вид насечки; б – увеличенное изображение насечки [13]
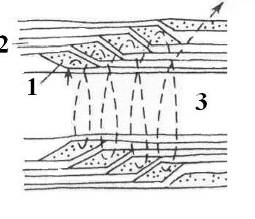
Рис. 4. Схема цитоплазматического канала (насечки) (1) в миелиновой оболочке (2) аксона. 3 – аксоплазма [13].
Краевая структура миелиновых оболочек в области перехватов Ранвье образует катушки из спиралей паранодальных петлей длиной порядка 1 мкм, сообщающихся с аксоплазмой через специальные окна. Если эти образования рассматривать как катушки индуктивности (Рис 5 - 7), то они будут играть существенную роль в сальтаторном механизме проводимости аксона.
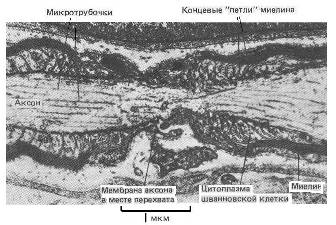
Рис. 5. Электронная фотография перехвата Ранвье, периферийного нерва [17].
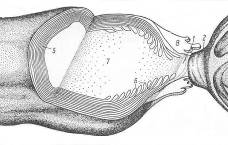
Рис. 6. Схема строения перехвата Ранвье. 1 – щель перехвата; 2 – мякотный конус; 5 – компактный миелин; 6 – расщепление основных плотных линий в области перехвата; 7 – осевой цилиндр (аксоплазма); 8 – цитоплазма шванновской клетки [13].
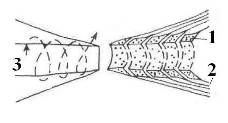
Рис. 7. Схема краевых спиралей петлей миелиновой оболочки в области перехвата Ранвье. 1 – цитоплазма; 2 – миелин; 3 – аксоплазма [13].
Различия электродинамических свойств аксоплазмы, мембраны и межклеточной жидкости, обусловленные различием их ионно-молекулярного состава и структуры, должны наложить свой отпечаток на механизм генерации ПД. Стимул, запускающий перезарядку мембраны, может быть как физической, так и химической природы, а само перераспределение зарядов может в той или иной пропорции сочетать перенос ионов через мембрану и их адсорбцию на ее поверхностях [14]. С ионными токами перезарядки поверхности мембраны аксона будут связаны импульсные токи смещения в паранодальных петлях и спиральных каналах насечек, что позволяет их уподобить магнитным диполям [9]. С кинетикой нарастания и последующей релаксации мембранного потенциала коррелирует кинетика ионных токов и токов смещения в аксоне, мембране и паранодальных петлях миелиновой оболочки. Поскольку фаза нарастания ПД длится около 0,1 – 0,2 мс, а время релаксации мембранного потенциала порядка ~1 мс [15], то и импульсные токи смещения, соответствующие фазе нарастания будут на порядок больше токов релаксации. Изменение заряда на внутренней стороне мембраны аксона в области перехвата Ранвье порождает волну поляризации или ток смещения в паранодальной области миелинового сегмента [16]. Величина данного возмущения будет экспоненциально затухать с расстоянием [17], а скорость распространения не превысит скорости движения ПД в немиелизированном нерве (порядка 1 м/с). Наличие окон связи паранодальных петель с аксоплазмой [16] обеспечивает преобразование волны поляризации в кольцевой ток смещения в спиралях петель.
Таким образом, генерирование ПД в перехвате Ранвье сопряжено с индуцированием и излучением-отшнуровкой вихревых ЭМ-квантов, метрику которых моделируют ЭМ-вихри в) и с) на Рис. 1. Возможно, что именно в этом и заключается главная функция концевых катушек миелиновых оболочек и спиралей насечек. Направление вектора плотности потока ЭМ-энергии (вектор Пойтинга) будет определяться знаком спирали. Данный фактор хиральности нейрона обеспечит односторонность распространения ЭМ-кванта, а значит, и ПД по миелизированному нерву. При достижении ЭМ-кванта со скоростью V (5) концевой катушки миелинового сегмента он может сыграть роль стимула для генерации ПД в следующем перехвате Ранвье. В данной модели сальтаторной проводимости нейрона скорость движения спайка будет лимитироваться процессом возбуждения тока в концевых катушках, время которого порядка 10–6 c (1 мкм : 1 м/с). При этом средняя скорость передачи ПД с одного конца миелинового сегмента на другой при его длине порядка 100 мкм и определит скорость сальтаторного механизма проводимости ~100 м/с.
Используя значение разности потенциалов, отвечающую ПД типичного нейрона (U ~ 70 мВ [15]), оценим величину электрической энергии, которая затрачивается на возбуждение ПД в перехвате Ранвье при сальтаторном механизме проводимости нейрона. Для этого представим перехват в виде цилиндрического конденсатора, обкладки которого образованы из мембраны нейрона и длина равна длине перехвата (f). Изменение энергии конденсатора (W), можно оценить по формуле:
W = (U2C)/2 . (8)
Величина С для цилиндрического конденсатора с расстоянием между обкладками (d) и радиусом внутреннего цилиндра (R) при условии d « R будет равна
С = (2peoe f)/[ln(1+d/R)] » (2peoe fR)/d
а величина
W = (U2 peoe fR)/d (9)
Подставим в (9) такие значения для нерва с R = 5 мкм [17]: U ~ 0,07В; eo = 8,85 ·10–12 Ф/м; e ~ 5; f ~ 10–7 м; d ~ 10–8 м , получим
W ~ 5·10–17 Дж или 3·107 Дж/моль. (10)
Такая же величина W получится, если подставить в (8) значение С = 10–2 Ф/м2 [17] при тех же параметрах перехвата Ранвье и величине U. Величина (10) сравнима с энергией, выделяемой при окислении ~10 молекул глюкозы и при гидролизе ~103 молекул АТФ.
Известно [12, 17], что при гидролизе одной молекулы АТФ через мембрану проходят ~3 иона Na+ в обмен на два иона К+, а при возбуждении ПД плотность потока ионов Na+ через мембрану перехвата составляет JNa ~ 4·103 ионов/мкм2. Тогда число вошедших в аксон ионов Na+ будет равно JNa (2pRf ) ~ 104, им соответствует ~3·103 молекул АТФ, суммарная энергия которых по порядку величины согласуется с (10). При концентрации АТФ в аксоплазме аксона кальмара ~1 ммоль на 1 кг Н2О [12], общее число молекул АТФ в цилиндре перехвата Ранвье (радиуса 5 мкм и длиной 1 мкм) будет равно ~ 4 ·107 молекул. Следовательно, величина W составит только 0,01% от полного энергетического ресурса перехвата Ранвье.
Очевидно, что энергия ЭМ-кванта, играющего роль стимула генерации ПД в перехвате Ранвье будет на один, два порядка меньше величины W. Например, за верхний предел энергии ЭМ-кванта можно взять энергию фотона с длиной волны 600 нм (4 ·10–19 Дж), которой достаточно, чтобы возбудить сигнал в рецепторной клетке сетчатки глаза [15].
2. 3. Нейросети
Подчинение нейрофизики закону ЭМИ можно формализовать, введя в эквивалентную электрическую схему мембраны нерва с миелиновой оболочкой вместе с конденсатором еще катушку индуктивности (Рис. 8). Такая модификация эквивалентной схемы, преобразуя ее в колебательный контур, существенно расширяет диапазон электрофизических свойств нейрона.
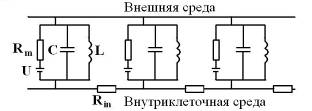
Рис. 8. Модифицированная электрическая схема мембраны нерва. Rm , U – ионный канал; С – емкость мембраны; L – индуктивность глиальных миелиновых спиралей мембраны; Rin – сопротивление аксоплазмы.
Помимо этого, введение катушек индуктивности в электрическую схему мембраны нерва позволяет смоделировать фактор хиральности нейрона и связать его с механизмом дифференциации нервных сигналов на возбуждающие и тормозящие. Сочетание фактора хиральности с биохимическим фактором (синаптические связи) наделяет логический элемент нейронных сетей возможностью кодировать сигналы “да” и “нет” (Рис. 9).
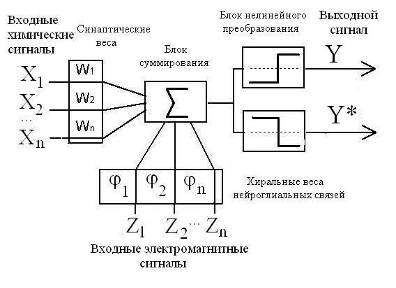
Рис. 9. Модифицированная функциональная схема формального нейрона [18]. Хn – биохимические, Zn – электрофизические факторы активности нейрона; Y (“да”), Y* (“нет”) – аналоги возбуждающего и тормозящего сигналов.
Кроме того, генерируемые в нейросети ЭМ-кванты или квазифотоны можно объединить в динамичную квантовую систему (Бозе-газ) и представить мозг процессором, элементной базой которого служит вся совокупность многоуровневой иерархии нейрон-нейронных и нейроглиальных связей. При этом оперативность квантового уровня организации нейросети будет лимитироваться величиной V (5) и время передачи и обработки сигнала в масштабе нейросети от 1 мкм до 10 см будет меняться в диапазоне от 10–15 до 10–10 с. Первое значение сопоставимо с временем жизни синглетного электронно-возбужденного состояния молекулы (оптический квазифотон), а второе с характерным временем жизни тетраэдрических кластеров воды.
2. 4. КАПСУЛИРОВАННЫЕ НЕРВНЫЕ ОКОНЧАНИЯ
2.4.1. Тельце Фатера Пачини
ЭМИ можно привлечь и для объяснения механизма генерации электрических импульсов в капсулированных нервных окончаниях. Наиболее важным для физики мозга представителем такого рода окончаний является тельце Фатера-Пачини (ТФП) (Рис. 10). ТФП в изобилии присутствуют в подкожном слое ладоней и стоп, в женских гениталиях и в соединительных тканях внутренних органов [19].
Рис. 10. Капсулированное окончание нерва – тельце Фатера-Пачини [19]. Линейные размеры достигают 1-2 мм.
Очевидно, что ТФП помимо сенсорной функции могут акцептировать гравитационную и геомагнитную энергии и одновременно исполнять роль генераторов ЭМ-квантов. Предполагают [19], что в основе механизма генерации ПД в ТФП лежит биохимический отклик ТФП на его механическую деформацию. Однако, изоморфизм ТФП и сложных электромагнитных устройств, имеющих на стержне две вложенных одна в другую катушек индуктивности, позволяет предположить участие ЭМИ в механизме генерации ПД. Деформация ТФП, будучи сопряжена с изменениями его индукционных или емкостных характеристик, может приводить к возбуждению электромагнитных импульсов, стимулирующих генерацию ПД. В этом случае снижение величин L и С в соответствие с формулой (6) должно привести к возрастанию частоты генерации стимулов (w), а значит, и частоты следования ПД, что и наблюдается на опыте [19]. Отметим, что при отсутствии внешней деформации ТФП их фоновую активность в качестве “генераторов” квазифотонов может обеспечивать ритмическая деформация клетчатки вокруг ТФП, отвечающая пульсации кровеносной системы. Не исключено также, что LC-контур в структуре ТФП при движениях рук и ног может резонансно поглощать энергию геомагнитного поля.
2.4.2. Глаз
Глаз можно считать интегральным капсулированным окончанием большого числа аксонов зрительного нерва. Его основная функция – преобразование фронта фотонов видимого диапазона в сложную пространственно-временную мозаику ПД и квазифотонов. Электрическая энергия (импульс) поглощенного сетчаткой фотона стимулирует генерацию в ней ПД и частично преобразуется в импульс спайка зрительного нерва. В такой роли выступают около 10% от попадающих в глаз фотонов, остальные 90% поглощаются оптическими средами глаза [19]. При поглощении фотонов, как в сетчатке, так в других элементах глаза высока вероятность генерации состояний с внутри- и межмолекулярным переносом заряда в донорно-акцепторых фрагментах (D+d–A–d). Такие метастабильные состояния называются экситонами. Кинетика процесса релаксации экситонов в сетчатке коррелирует с кинетикой генерации ПД. Достаточно большое время жизни и высокая фотостационарная концентрация этих состояний обусловливают дипольную поляризацию поверхности сетчатки [20]. Перемещение зарядов сетчатки при движении глаз индуцирует вихревые магнитные поля (ЭМ-вихрь), максимальная плотность энергии которых достигается в лобных и височных долях, а также в пазухах черепа (верхнечелюстных, клиновидных, лобных) (Рис. 11). Известно [11], что в данных областях локализованы функции внимания и самосознания, поэтому ЭМ-вихри глаз могут принимать прямое участие в их активации. С учетом этого предположим, что глаза и их нервная система наряду со своей сенсорной функцией играют доминирующую роль в физике когнитивных функций мозга. Именно поэтому при усиленной умственной работе, даже не связанной с чтением, сильно устают глазные мышцы, что провоцирует развитие специфического рисунка морщин вокруг глаз. Отметим, что при врожденном поражении отделов ЦНС, ответственных за формирование наглядных представлений внешнего мира (“центральная врожденная слепота”), ребенок обречен остаться идиотом.
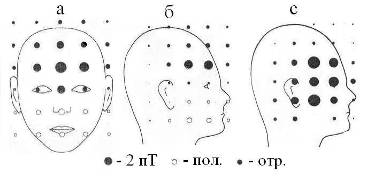
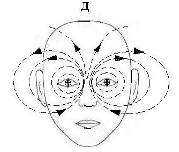
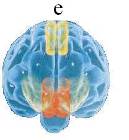
Рис. 11. Распределение магнитной индукции при различных движениях глаз (а, б, с) [20, 21] и схема потоков магнитной индукции во фронтальной проекции (д). Пол. – магнитное поле направлено внутрь, отр. – наружу объекта. Величина В-поля пропорциональна радиусу кружка. А), б) – горизонтальное движение глаз справа налево в пределах угла в 55о; с) – движение глаз снизу вверх. На Рис 11е показаны области мозга (вид спереди), ответственные за самосознание: красным цветом выделена медиальная префронтальная кора (связывает самоощущения и память о себе); желтым – предклинье (активация ретроспективной памяти о себе) [11].
Геометрия вихревых B- и D-полей глаз задается траекториями зарядов сетчатки и мышц при движении глаз в вертикальном и горизонтальном направлениях. Локализация максимального значения В-поля при горизонтальном перемещении глаз в срединной точке (Рис. 11а) указывает на суммирование в этой точке В-полей от обоих глаз. Такое возможно при условии, если метрики вихрей индуцируемых правым и левым глазам зеркально симметричны. Не исключено, что хиральность электромагнитного стимула, а значит, и знаки миелиновых спиралей нервов правого и левого глаза противоположны. Можно представить, что ЭМ-вихри индуцируют в ликворе продольной щели между полушариями зеркально симметричные пары ЭФ, их слияние, в принципе, может давать квазифотон и такой механизм генерации ЭМ-квантов, очевидно, лежит в основе экзотермического процесса рекомбинации двух разноименных электрических зарядов. Очевидно, что организующее действие ЭМ-вихрей глаз в процессе формирования самосознания ребенка обусловливает образование в медиальной префронтальной области коры нейронов уникальной формы, называемых клетками-веретенами [11, 15]. Аналогичные реакции слияния-рекомбинации ЭФ, индуцируемых в правом и левом полушарии, могут идти также и в ликворе третьего и четвертого желудочков, принимая активную роль в их биоэнергетике. Упрощенная схема фронтальной проекции D-, B-вихрей глаз показана на Рис 11д. Крестик в центре глаза обозначает уходящий в плоскость рисунка спайк зрительного нерва. Данная схема совпадает также с распределением силовых линий магнитного диполя, ориентированного по линии носа. Это согласуется также с тем фактом, что пористые кости стенок носа, клиновидной пазухи и решетчатой кости имеют высокое значение остаточной намагниченности [30].
В подтверждение важной роли глаз в когнитивной физике мозга говорит наличие сложных взаимоотношений между энергетикой глаз и базовым ритмом электрофизики мозга (альфа-ритмом):
- альфа-ритм имеют только высшие млекопитающие [22] и он устанавливается синхронно с половым созреванием, после чего он не фиксируется в лобных долях [15];
- альфа-ритм, как и бета-ритм, может локализоваться обособленно в правом- или левом полушарии мозга [23];
- альфа-ритм исчезает при потере сознания и открывании глаз, однако у слепых он либо плохо выражен, либо отсутствует [15];
- частоту альфа-ритма (~10 Гц) соотносят с частотой стоячей ЭМ-волны в сферическом резонаторе, который образует поверхность Земли и ее ионосфера [24].
Стекловидное тело глаза в фоновом режиме может конденсировать ЭМ-энергию внешней среды и, возможно, энергию солнечного нейтрино [1, 6], напрямую питая этой энергией мозг. В пользу данного предположения свидетельствуют данные:
– быстрое движение глаз в фазе парадоксального сна (фаза-БДГ) сопряжено с интенсификацией физики мозга [15];
– образование специфического пятипальцевого рельефа на поверхности глазницы, обращенной к мозгу и искривление линии носа [25];
– мышцы глазного яблока спонтанно подергиваются с частотой 20 – 150 Гц (микросаккады, тремор) [15];
– граничащий со стекловидным телом слой ганглиозных клеток сетчатки в темноте и при закрытых глазах проявляют фоновую активность с частотой от 1 до ~20 импульсов в секунду [26];
- воздействие на закрытые глаза механического давления и импульсного магнитного поля инициирует “видение” белого света (фосфены) [15, 27]
- увеличение интенсивности света ведет к возрастанию частоты генерации ПД в зрительном нерве [15];
- хрусталик и стекловидное тело глаза оптически активны [28, 29];
- гликолиз глюкозы в стекловидном теле дает наряду с АТФ еще хиральную молочную кислоту [15];
- характерное время гидродинамики глаза составляет ~900 с, за это время обновляется половина жидкости стекловидного тела [15].
Онтогенез асимметрии зрения, обоняния, слуха, лица (искривление носа) и половых органов синхронизован с процессом стабилизации частоты альфа-ритма [31] и за 12 – 13 лет повторяет этап филогенеза, соответствующий прямохождению. Причем уже к двум годам, когда ребенок начинает самостоятельно ходить, в генезис асимметрии мозга включается энергетика ТФП стоп и физика половых органов, гендерные особенности которой накладывают свой отпечаток на топологию и функции мозга мужчины и женщины [32].
3. КВАЗИФОТОН
3.1. Типы квазифотонов
Для описания свойств различных конденсированных сред широко используют понятие квазичастица [33]. Поскольку содержание воды в мозгу достигает ~75% [15], его можно считать высококонцентрированным коллоидным раствором. Для описания механизмов энергоинформационных процессов, лежащих в основе физики мозга, удобно использовать понятие квазифотона, как обобщение ЭМ-кванта. Таким образом, квазифотон является носителем избыточной энергии электромагнитного поля, локализованной на электроне или на системе электронов той или иной упорядоченной атомно-молекулярной структуры. Предшественником квазифотона могут быть фотон или ЭМ-квант, в случае их поглощения системой. В зависимости от энергии фотона и электронной структуры системы метрика, время жизни и судьба квазифотона варьируются в широких пределах. Физика квазифотонов генетически наследует законы атомно-молекулярной спектроскопии и свойства возбужденных состояний молекул различных типов (электронные, колебательные, трансляционные, вращательные) [34]. Смешанным электронно-ядерным конфигурациям возбужденных состояний будут отвечать вращательные и колебательные квазифотоны, а чисто электронным возбужденным состояниям – оптические квазифотоны. Примерами оптического квазифотона служат, ЭМ-стимул генерирующий ПД, экситон или электронно-возбужденное состояние молекулы. Колебательный квазифотон в упругой связанной структуре подобен фонону. Вращательный квазифотон в системе связанных ядерных или электронных спинов можно отождествить с магноном [33]. Квазифотоны могут быть свободными и связанными в зависимости от свойств среды и механизма взаимодействия ее элементов. Таким образом, метрико-динамические характеристики квазифотонов будут определяться типом химических связей и видом межмолекулярных взаимодействий, которые определяют степень упорядоченности среды.
Энергия квазифотонов различных типов меняется в широком диапазоне, верхней границей которого можно считать энергию квазифотона стимулирующего генерацию ПД в перехвате Ранвье (~10–19 Дж). За низший предел энергии квазифотона можно принять энергию вихревого ЭМ-поля, генерируемого движением глаз. Плотность данной энергии при В ~ 4 пТ имеет порядок:
Е = В2/(2mоm) ~ 10–23 Дж/см3 или ~0,01 кДж/моль в см3. (11)
Степень влияния данного поля на магнитно-восприимчивые микро и макро структуры и среды мозга будет определяться величиной плотности энергии:
Е = (М В)/2,
где М - удельная намагниченность (удельная плотность магнитных моментов - m), равная:
М = S m.
В случае кольцевых токов любой природы (J) m = J DS, где DS – площадь поверхности, охватываемой током. Причем поляризационный эффект магнитного поля может усиливаться под влиянием теплового движения частиц среды [36].
Сравнима с величиной (11) энергия теплового эффекта от светового раздражения глаз крысы, который проявляется повышением локальной температуры зрительной коры мозга на ~0,06о С [11]. Величина энергии квазифотона отвечающего данному кванту тепловой энергии составит ~10–24 Дж или ~10–3 кДж/моль. В диапазон 10–3 – 102 кДж/моль попадает энергия биогенного МКВ-излучения (l = 100 – 1 мм, Е = 10–3 – 0,1 кДж/моль) [37]; в том числе и энергия резонансных частот воды (l ~ 6 мм, Е = 0,02 кДж/моль) [38]. Известно [3], что энергия активации процессов ассоциирования сахаров и квантов биогенного МКВ-излучения на один-два порядка меньше тепловой энергии и сравнима по порядку величины с (11). Отсюда следует, что в процессах самоорганизации жидких сред мозга ключевую роль играют квазифотоны вращательного типа и физика лобно-височных долей, ответственная за когнитивные функции мозга, непосредственно связана с электрофизикой глаз.
3.2. Метрика квазифотона
Элементарной структурной ячейкой жидкой воды является динамический тетраэдр, образованный из четырех молекул воды, связанных между собой водородными связями. Пятая молекула воды или соразмерная с ней молекула или атом могут находиться в центре тетраэдра, тогда он называется центрированным тетраэдром (Рис 13). Благодаря водородным связям, вода эффективно взаимодействует с растворенными молекулами, расширяя тем самым спектр их физико-химических свойств. Данная особенность водных коллоидов и гелей особенно важна для физики мозга, поскольку его межклеточные объемы, как правило, сравнимы с размерами биомолекул, клеток и органелл [35].
В силу этого следует предполагать существенное влияние эпитаксиального эффекта на процессы, регулирующие межнейронные и нейроглиальные взаимодействия. Известно, например, что в химических реакциях, протекающих в оптически активной среде или на поверхности кварца, возрастает выход хиральных продуктов. Увеличению эпитаксиального эффекта мембран и стенок различных органов, помимо посредничества воды, очевидно, способствуют связанные или адсорбированные поверхностью полипептидные и полисахаридные цепочки, а также микроворсинки (Рис 12) [15, 17]. Эпитаксиальный эффект и присутствие хиральных сахаров сказывается на кинетике обратимой адсорбции ионов и нейромедиаторов на поверхностях мембран нейронов как в перехватах Ранвье, так и в синапсах [14].
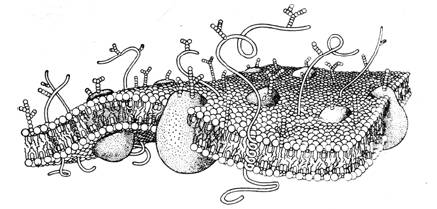
Рис. 12. Схема мембраны и выходящих из нее полисахаридных и полипептидных цепочек
Метаболиты, имеющие заряд, диполь, механический или магнитный моменты, а также хиральность, влияя на электродинамическую постоянную (em) среды, метрику и динамику надмолекулярных структур, могут в широких пределах менять кооперативные свойства растворов, эффективность генерации и механизм движения квазифотонов. Это относится, прежде всего, к ионам (Na+, K+, Cl–, Р3+) (Таблица 1) и к молекулам, играющим роль переносчиков, акцепторов и преобразователей квазифотонов (кислород, углекислый газ, вода, сахара, АТФ, нейромедиаторы, гормоны, ферменты).
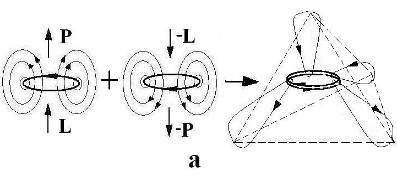
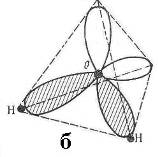
Рис. 13. Схема слияния двух зеркально симметричных подвижных ЭФ (n/g-пар) в покоящийся квазифотон с тетраэдрической метрикой (а) и схема электронных орбиталей молекулы воды (б)
Основным механизмом движения квазифотонов будет их резонансное поглощение и переизлучение молекулами среды, метаболитами и надмолекулярными структурами. Главным элементом трехмерной метрики жидкой среды и большинства органических метаболитов служит тетраэдр, электронно-ядерной матрицей которого является sp3-гибридизированная система электронных орбиталей атомов углерода, азота и кислорода. Следовательно, квазифотон, локализованный на том или ином метаболите, с наибольшей вероятностью будет иметь метрику изоморфную геометрии sp3-гибридизации.
Используя представление о n/g-парах, покоящуюся ЭФ или локализованный квазифотон с тетраэдрической метрикой можно получить по схеме, показанной на Рис.13. Правила комбинирования и конденсации ЭФ (n/g-пар) [6] позволяют моделировать и рассчитывать метрику квазифотонов различных типов, в том числе изоморфных метрике sp- и sp2-гибридизированных атомных орбиталей. Энергия квазифотонов, связанных с p-электронами, будет меньше энергии квазифотонов, отвечающих колебательно-вращательным возбуждениям атомов или деформациям s-скелета. Низшие колебательные уровни молекулы углекислого газа (О=С=О), имея энергию от 10–21 до 10–20 Дж, могут заселяться за счет поглощения тепловых квантов (kT). Специфика расположения уровней допускает их инверсную заселенность, что позволяет использовать углекислый газ в качестве активной среды лазера (l ~ 10 мкм). В жидких средах предрасположенных к самоорганизации молекула СО2 может быть донором колебательных квазифотонов для молекул с карбоксильной группой (–НСО). Аналогично, молекулы с ароматическими циклами будут акцепторами квазифотонов, отвечающих конформационным колебаниям изоморфных им насыщенных углеродных циклов и гетероциклов. Высокая активность, например, стероидных гормонов производных холестерола, имеющих конденсированные гексановые цикла, может быть обусловлена насыщенностью их молекул квазифотонами с sp3-метрикой (Рис 13). При этом изоморфные фрагментам гормонов ароматические молекулы (бензол, антрацен, пирен), эффективно дезактивируя гормоны и искажая их метаболические функции, могут инициировать канцерогенез [39].
Таблица 1.
Ядерно-электронные характеристики элементов
Элемент |
Характеристики ядра |
Основное электр. состояние Атома |
Ионный радиус (нм) |
|||
Спин (I) |
Магнитный момент (ґ mн) |
Магнитная воспри-имчивость (C13 = 1,00) |
Квадру-польный момент (Qґ1028, м2) |
|||
Натрий 11Na23 |
3/2 |
2,22 |
525,0 |
0,12 |
[Ne]3s1 |
0,098 |
Калий 19 K39 |
3/2 |
0,39 |
2,7 |
0,055 |
[Ar]4s1 |
0,133 |
Хлор 17 Cl3517 Cl37 |
3/2 |
0,82 0,68 |
20,2 3,8 |
– 0,08 – 0,06 |
[Ne]3s22р5 |
0,182 |
Фосфор 15 Р3115 Р32 (14 дней) |
1/2 1 |
1,13 –0,25 |
377 |
– – |
[Ne]3s23р2 |
0,044 |
3.3. Метаболические квазифотоны
Для унификации языка биоэнергетики соотнесем с величинами энергий химических связей, колебательно-вращательных и тепловых движений атомов и молекул энергию квазифотонов соответствующей метрики. Дееспособность мозга обеспечивает энергия ферментативные реакции окисления глюкозы в митохондриях и анаэробного ее гликолиза в глазном яблоке. В этих реакциях ЭМ-энергия химических связей глюкозы и кислорода трансформируется в энергию макроэргических связей АТФ, которая в последующих реакциях гидролиза АТФ преобразуются в кинетическую и колебательно-вращательную энергию метаболитов и молекул среды. Химическая активность этих молекул реализуется затем через действия их энергии возбуждения, которую и моделируют квазифотоны соответствующей энергии и метрики.
Суммарный энергетический эффект всех стадий ферментативной реакции окисления глюкозы в митохондриях имеет своим пределом тепловой эффект реакции горения глюкозы в атмосфере кислорода:
С6Н12О6 + 6 О2 > 6 СО2 + 6 Н2О + 2800 (кДж/моль). (12)
Реакция окисления глюкозы в митохондриях сопряжена с реакцией синтеза АТФ, при этом на одну молекулу глюкозы приходится 38 молекул АТФ [40]. При анаэробном гликолизе глюкозы образуются только две молекулы АТФ и две молекулы хиральной молочной кислоты, которые, очевидно, вносят свой вклад в хиральность энергетики глаз и мозга. Максимальный выход метаболической энергии даст гидролиз 38 молекул АТФ по схемам:
АТФ > АДФ + Р~P + 36 (кДж/моль)
Р~Р > Р + Р + 33,4 (кДж/моль).
Полная энергия макроэргических связей 38 молекул АТФ равна 2640 кДж/моль, что составляет ~95% от предельного значения энергии сгорания одной молекулы глюкозы. Это говорит о высокой эффективности ферментативных реакций трансформации квазифотонов, соответствующих s-связям С-С, С-О-С, С-Н глюкозы в квазифотоны, локализованные на двух макроэргических связях Р~О- в АТФ.
Предположим, что квазифотоны равновероятно распределяются по связям продуктов реакций окисления глюкозы и гидролиза АТФ, тогда предельные значения энергий квазифотонов, отвечающих данным реакциям будут равны 1/12 и 1/152 от теплового эффекта реакции (12), равного 4,5 ·10–18 Дж, то есть ~3 ·10–19 и ~3 ·10–20 Дж, соответственно. Если к этим квазифотонам применить универсальное соотношение между энергией и характерным размером (r) дискретного элемента материи (n/g-пара, элементарная частица) [6]:
Е ~ hc/r , (13)
то для квазифотона, действующего в виде кванта метаболической энергии, получим радиус ~1 мкм, сравнимый с радиусом аксона.
Метаболические квазифотоны могут принимать активное участие в ферментативном синтезе белков и нуклеиновых кислот, а также в репликации и транскрипции ДНК. Можно представить участие квазифотонов в расплетении двойной спирали ДНК следующим образом. В области репликативной вилки сахарофосфатный остов цепи ДНК резонансно поглощает метаболические квазифотоны колебательного типа. Возрастает упругость цепей, что и приводит к разрыву водородных связей между ними. Учитывая, что на два сахарофосфатных звена спирали ДНК приходится одна водородная связь и ее энергия равна ~19 кДж/моль (3 10–20 Дж), получится, что для ее разрыва достаточно поглощения цепью ДНК одного метаболического квазифотона.
Присутствие изоморфных аминокислотных фрагментов в пептидных цепях белка и в структуре нейромедиаторов (глицин, ацетилхолин, глутаминовая кислота, дофамин, серотонин и др.) позволяет предложить резонансный механизм передачи квазифотона колебательного типа при контакте нейромедиатора с рецептором. Из-за наличия в структурах медиаторов электроно-, протонодонорных и акцепторных групп их основное электронное состояние характеризуется внутримолекулярным переносом заряда Д+d-С-А-d. Здесь Д – аминогруппы, метоксигруппа, бензольное кольцо и А – карбонильная и гидроксильные группы, а С – цепочка из s-связей. Этот фактор и предрасположенность медиаторов к образованию водородных связей лежат в основе их физической и химической сорбции на рецепторах постсинаптических мембран. Рецептор, принимая или отдавая квазифотон при контакте с нейромедиатором, меняет свою конформацию, запирая или открывая при этом кальциевый канал мембраны.
4. ТЕРМОДИНАМИКА МОЗГА
Мозг в целом можно считать реакционной термодинамической системой, находящейся в стационарном состоянии. Приток энергии и сброс избыточного тепла мозгом сбалансированы в узком диапазоне температур от ~37о (центр мозга) до ~36оС (кора мозга) [15]. Этот градиент температуры, будучи обусловлен более низкой температурой внешней среды, может играть существенную роль в ориентировании тепловых потоков внутри мозга. Аналогичный градиент температуры наблюдается и для тела, она имеет максимум в прямой кишке, а минимум в поверхностном слое клетчатки и мужских яичках [15]. Диапазон оптимальной температуры метаболизма находится в пределах значений температур, для которых изобарная теплоемкость чистой воды имеет минимум [41]. Особенности термодинамики фазовых переходов водных растворов в процессе филогенеза легли в основу механизма адаптации живых систем, которая, по сути, представляет собой изоэнергетические переходы или переходы с энергией активации порядка kDT (при DT ~ 0,1 – 1 К) между состояниями разной степени упорядоченности белковых молекул или однородных, молекулярно-клеточных ансамблей. Снижение энтропийной составляющей внутренней энергии живой системы сопряжено с резонансным поглощением ею кванта внешней ЭМ- или нейтринной энергии, который она преобразует в активный метаболический квазифотон [3]. Жидкостная среда обеспечивает отвод кванта тепловой энергии (энтропии) за границы системы, а действием квазифотона реализуется функция той или иной структуры мозга, включающей в себя упорядоченную подсистему. Энергия, выделяемая или поглощаемая при таких переходах, может оказаться намного порядков меньше kT. В неравновесных условиях колебания отдельных макромолекул могут синхронизироваться, в частности, посредством электромагнитного поля [36].
Таким образом, термодинамика мозга сочетает равновесно-стационарную термодинамику метаболизма и неравновесную термодинамику нейросети, “рабочего телом” которой является Бозе-газ квазифотонов. Соответственно, внутренняя энергия U мозга как функция его состояния будет зависеть в общем случае от температуры (или энтропии S), от тензора деформаций G, зависящего от внутричерепного давления, от магнитного момента M отдельных метаболитов и макроструктур, от суммарного момента количества движения ядер и атомов L и от поляризация среды Р. Следовательно, полный дифференциал внутренней энергии U = U (S, G, M, L, P) будет иметь вид:
dU є TdS – ПdG + ВdM + DdP + FdJ, (14)
где Т – абсолютная температура системы; П – тензор давлений; F – вектор ориентационной поляризации системы спинов или моментов импульса [42]. В выражении (14) член FdL характеризует работу, связанную с ориентационной поляризацией системы ядерных спинов или моментов импульса атомов и молекул (подобно тому, как члены DdP и BdM определяют работу, связанную с поляризацией и намагничиванием системы).
Ориентационные и поляризационные эффекты существенную роль играют в инициации фазовых переходов в однородных газовых и жидкостных системах мозга и организма. Высокую чувствительность данных систем к параметрам входящим в (14) обеспечивает хиральность метаболитов (в основном сахаров [3, 29]) и физико-химические особенности молекулярной и жидкой воды. Такие системы формируются в следующих структурах и средах организма и мозга:
– желудок, матка, трахея, черепно-лицевые пазухи, полость эпифиза;
– оболочки и желудочки мозга, венозные синусы, глазное яблоко;
– кровеносная и лимфатические системы;
– паренхима органов (легкие, печень, селезенка, яички, женская грудь);
– подкожная клетчатка, соединительная и костная ткань.
Все перечисленные системы в норме функционируют в двух режимах – стационарном (квазиравновесном) и неравновесном. Первый характерен для бодрствующего состояния организма и мозга не занятого мыслительной работой, а второй режим соответствует состоянию сна или творческой работе. В первом режиме обмен энергией со средой происходит непрерывно, а во втором – квантуется. Механизм акцепции кванта внешней энергии в фазовом переходе кооперативной системы иллюстрируют процессы конденсации паров воды в точке росы и квантовой Бозе-конденсации.
Акцептированию квантов энергии МКВ-диапазона или нейтринной энергии в указанных средах способствует снижение температуры организма во сне на ~1К, а также пониженные температуры стекловидного тела глаз, периферийной (депонированной) крови и яичек. В акцепции хиральных квантов нейтринной энергии большую роль играют сахара, содержание которых в крови возрастает в утренние часы до восхода солнца. В это время нейтринная составляющая солнечного излучения отфильтровывается от ЭМ-излучения поверхностным сегментом коры земного шара (Рис. 18). Важную роль в акцепции энергии стекловидным глазом играет полисахарид - гиалуроновая кислота [3, 29]. Почти половина всей гиалуроновой кислоты организма человека сосредоточено в его коже, где она располагается в соединительной ткани дермы между волокнами коллагена и эластина, а также в клетках рогового слоя корнеоцитах. В дерме содержится 70% воды, что составляет ~20% всей воды организма [15]. Поглощаемая организмом энергия, конденсируясь на метаболитах в составе жидких сред (кровь, спинномозговая жидкость), передается в мозг по нейрогуморальным и воздушным каналам (из легких).
Поглощение внешнего ЭМ-кванта и формирование квазифотона из энергоформ в общем случае подчиняется фрактально-резонансному механизму и принципу изоэнергетичности. С учетом (13) принцип изоэнергетичности для резонансных взаимодействий и фазовых переходов в кооперативных системах можно выразить соотношением [6]:
hC/r = N (hC/R), (15)
здесь r характеризует метрику квазифотона, а R – энергоформы и r = R/N; число N принимает любые значения меньшие числа Авогадро при конденсации ЭФ в квазифотоны и достигает числа Авогадро при участии ЭФ в слабых взаимодействиях [39].
5. ФИЗИКА ОРГАНИЗАЦИИ МОЗГА
5.1. Функциональная иерархия мозга
Функциональная иерархия мозга человека строится на физических свойствах следующих его структур: неокортекс, базальные ядра, лимбический мозг, таламус, гипоталамус, гипофиз, эпифиз, ретикулярная формация и мозжечок. К отдельным элементам иерархии следует отнести жидкостные системы мозга (кровеносная и ликворная), а также весь комплекс внутричерепных нервных коммуникаций, выделив в нем мозолистое тело, зрительный нерв и лучистости таламуса.
5.1.1. Кора больших полушарий
В структуре коры различают поверхностные специализированные зоны и шесть слоев [15]. Самый верхний слой образуют горизонтально ориентированные апикальные дендриты пирамидных клеток и аксоны звездчатых клеток, которые обеспечивают внутрикорковые связи между соседними нейронами. Горизонтальной ориентации диполей нейронов данного слоя соответствует динамичное электрическое поле, которое может играть роль защитного ЭМ-экрана. Остальные пять горизонтов коры структурируются сначала в нейро-глиальные модули (диаметр ~ 100 – 150 мкм), а затем, в колонки диаметром до 1 мм и со средним числом нейронов ~100 шт [15]. Синхронизация электрической активности нейронов в модулях и колонках приводит к формированию в пучках отходящих от них аксонов залповых импульсов. Процессу суммирования ПД в залпы может предшествовать конденсация по (15) внешних ЭМ-квантов или квазифотонов стимулов ПД на отдельных нейронах в модулях, а затем в колонках. Например, при конденсации ~100 квазифотонов с характерным радиусом 100 мкм в залповом импульсе может образоваться квазифотон радиуса 1 мкм (энергия ~10–19 Дж), который выдаст колонка на сопряженную с ней структуру подкорки или инжектирует в ликвор.
Внешний слой коры вместе со слоем ликвора в подпаутинном пространстве мозга, очевидно, играют роль сферического ЭМ-фильтра или МКВ-резонатора (R ~ 7 – 10 см), поглощение которого имеет несколько максимумов и в том числе для длин волн внешних ЭМ-квантов (l) порядка радиуса сферы [37]. В тканях мозга длина волны МКВ-кванта снижается в ~7 раз, и глубина его проникновения оказывается одного порядка с l [37].
Можно предположить, что горизонтальное расслоение коры (Рис. 14) связано с дисперсией интенсивности поглощения веществом мозга биогенных МКВ-квантов. И при толщине коры ~2 – 5 мм ее расслоение на три миелиновых слоя (Рис. 14) может быть филогенетически обусловлено действием трех типов ЭМ-квантов с длиной волны от 1 см до 10 см. Предположив, что из ЭМ-квантов с l~ 1 см формируются квазифотоны-стимулы ПД, по формуле (15) можно оценить их число (N), исходя из радиуса квазифотона в 100 мкм. Оценка N дает величину ~ 1 см/100 мкм = 100, равную числу пирамидных нейронов в колонке.

Рис. 14. Микрофотография среза коры прокрашенного красителем, поглощаемым миелином.
5.1.2. Таламус, желудочки мозга
Геометрическим и энергоинформационным центром мозга является двудольный таламус, имеющий ось, ортогональную плоскости III-го желудочка (межталамическое сращение). Его веерные нервные связи с корой (лучистость, Рис. 15) при синхронизации своей электрической активности могут порождать макроскопический ЭМ-вихрь с магнитным вектором, направленным по оси продолговатого мозга.
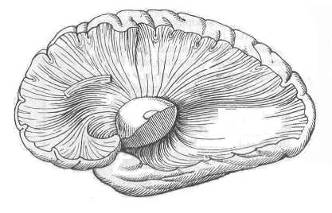
Рис. 15. Лучистости таламуса
Аналогичным образом, залповые импульсы в нервных волокнах гиппокампа будут генерировать ЭМ-вихрь с магнитным вектором параллельным оси межталамического сращения. Тонкий слой ликвора в желудочках мозга насыщенный метаболитами под влиянием эпитаксиальных эффектов и в условиях сна, очевидно, переходит в жидкокристаллическое состояние, приобретая свойство конденсора-синтезатора макроскопических квазифотонов мозга или внешних энергоформ (в том числе и нейтринных). Физико-химическая связь ликвора III-го желудочка с железами эпифизом и гипофизом обеспечивает их участие в механизмах адаптации, гомеостаза и размножения. Тела обеих желез погружены в ликвор цистерн мозга, а их горлышки-воронки контактируют с ликвором III-го желудочка (Рис. 16). Причем, если эпифиз участвует в формировании энергоинформационных кодов ликвора и венозной крови, то гипофиз только транслирует некоторые из них на информационный уровень кровеносной системы.
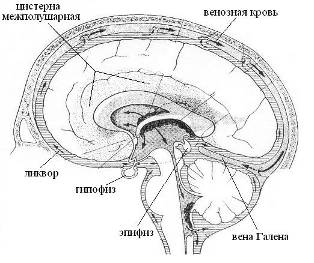
Рис. 16. Цистерны мозга (заштрихованы). Черные стрелки обозначают токи венозной крови
5.1.3. Эпифиз
В работах [1, 6] высказана гипотеза, что филогенез зрительной системы и эпифиза как третьего глаза обусловлен воздействием на биосферу излучения Солнца не только электромагнитной, но и нейтринной природы. Согласно стандартной модели Солнца доля нейтринной энергии составляет около 3% от ЭМ-энергии и поэтому для выявления ее эффектов в дневное время ошибка эксперимента или статистики не должна превышать ~1%. Поскольку нейтрино практически не поглощается косным веществом, оно легко проходит сквозь Землю, магнитное поле которой может только приводить к дисперсии или фокусировке падающего на нее потока нейтрино. Таким образом, можно ожидать существенный вклад нейтринной энергии в физику мозга в ночные часы и когда мозг пребывает состоянии сна. Очевидно, что сама потребность во сне филогенетически обусловлена нейтринным фактором, под влиянием которого в жидких средах организма и развились механизмы конденсации квантов биогенной энергии различной природы. Особенно большое значение данная энергетика имеет для развития эмбриона в чреве матери, а затем младенца грудного возраста. Помимо функциональных специфических особенностей мозга женщины [32] ее организм в период лактации интенсифицирует процесс акцепции нейтринной энергии с участием метаболитов, входящих в состав грудного молока. Возможно, что этой физической особенностью материнского молока и обусловлен быстрый темп развития и структуризации неокортекса у млекопитающих, как в филогенезе, так и в онтогенезе.
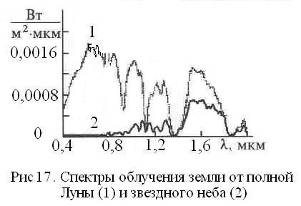
Нельзя также исключать и действие на живые системы в ночное время солнечных ЭМ-квантов, отраженных Луной и планетами (Меркурием, Венерой, Марсом) (Рис. 17). Интенсивность этих излучений, очевидно, сопоставима с интенсивностью потока нейтринной энергии, а их суммарное действие будет иметь максимальный эффект тогда, когда Марс и Венера находятся в противостоянии с Землей. Поскольку атмосфера Венеры состоит в основном из СО2, то спектр отраженного (переизлученного) ею света будут промодулирован колебательными частотами СО2, что обеспечит резонансное возбуждение молекул СО2 в земной атмосфере и в средах насыщенных СО2 (морские воды, литосфера, венозная кровь). В определенных космических ситуациях на биосферу в ночное время могут оказывать влияние и галактические излучения электромагнитной и нейтринной природы. Вся эта гамма физических факторов филогенеза и отразилась в особенностях физики и химии спящего мозга, о чем, в частности, свидетельствуют данные хронобиологии мозга (Рис. 18).
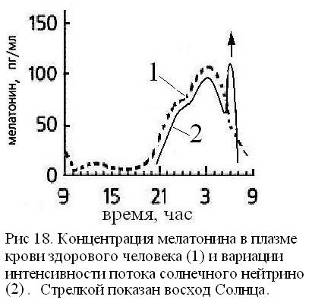
Одним из путей метаболизации нейтринной энергии может служить реакция синтеза в эпифизе нейрогормона мелатонина, играющего ключевую роль в процессе полового созревания и умственного развития, а также в формировании спектрально-энергетических характеристик поверхностного слоя кожи. Процесс синтеза мелатонина модулирован циркадными ритмами и подавляется ярким светом. Мелатонин выбрасывается в ликвор III-го желудочка и в венозную кровь, причем его максимальная концентрация в ликворе наблюдается около трех часов ночи, достигая на порядок большей величины, чем его содержание в крови [43]. Отметим, что в три часа ночи минимальное значение имеют артериальное давление и температура организма, что способствует процессу самоорганизации жидких сред и повышает тем самым эффективность акцептирования ими квантов биогенной энергии [3, 29]. В процессах акцепции и утилизации энергии помимо мелатонина участвуют хиральные сахара и фосфор. Их концентрации достигают максимума также в ранние утренние часы до восхода Солнца [15] (Рис. 18). Летом в это же время выпадает роса. Насыщение нейтринной энергией жидких сред интенсифицирует ферментативное окисления сахаров, что в сумме с повышением концентрации фосфора дает увеличение скорости синтеза АТФ. Таким образом, нейтринная энергия в ночные часы может исполнять роль хирального кинетического фактора биоэнергетики.
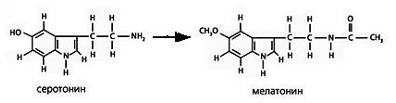
Рис. 19. Схемы структур серотонина и мелатонина
Филогенетическим и химическим предшественником мелатонина является нейрогормон серотонин (Рис. 19). Замена заместителей в кольце и при азоте при переходе от серотонина к мелатонину существенно меняет его физико-химические свойства. Благодаря -С=О группе мелатонин будет акцептировать квазифотоны от СО2 венозной крови. Кроме того, мелатонин может служить акцептором квантов энергии, которые конденсируются в стекловидном теле глаза и в ликворе в состоянии сна.
В метаболизации биогенной энергии вместе с мелатонином могут участвовать и другие нейрогормоны. В первую очередь это относится к дофамину, о чем свидетельствует, например, такой опыт. Прием в течении двух недель мотилиума (антагонист периферических дофаминовых рецепторов) и омепразола (ингибитор водородной помпы) наряду с положительным эффектом привел к снижению либидо, нарушению сна (регулярные пробуждения в три часа ночи) и усилению к этому времени аллергической реакции кожи (крапивницы). Синхронизация данных расстройств с кинетикой синтеза мелатонина (Рис. 18) позволяет связать их с гипертрофией гормональной функции мелатонина, причиной которой может быть концентрационный дисбаланс между мелатонином и дофамином.
Поскольку эпифиз полностью погружен в несжимаемый ликвор и рядом с ним находится крупный венозный узел, включающий большую мозговую вену Галена (Рис. 16), то объем эпифиза, а значит, объем и давление в его внутренней полости [44] будут пульсировать. При этом на этапе расширения на молекулах газа в объеме и на структурах внутренней поверхности полости будут конденсироваться кванты биогенной энергии, которые на этапе сжатия могут транслироваться вместе с метаболитами в паренхиму эпифиза, активизируя в ней клеточный метаболизм, а также в ликвор III-го желудочка.
5.2. Кинетические параметры физики мозга
Ритмика некоторых физических процессов в мозгу может быть обусловлена периодическими метаболическими реакциями. К таким процессам, очевидно, относится пульсация нейроглии – 12 секунд фаза напряжения и 240 с фаза расслабления, при этом меняется их объем, набухают и отбухают их отростки [45]. Колебания химической активности синапсов имеют постоянную времени порядка 100 мс, которая соответствует суммарной длительности возбуждающих и тормозных постсинаптических потенциалов ~30 и ~70 мс, соответственно. Учитывая доминирование циркадных ритмов в хронобиологии, можно полагать, что в основе механизма работы ритмоводителя мозга или сердца лежит связь электрической активности специальных нервных клеток с тем или иным периодическим явлением электромагнитной природы геофизического или космического масштаба [46]. Примерами таких явлений могут служить спонтанное реликтовое излучение, пульсации геомагнитного поля и его периодические возмущения Солнцем, Луной и другими планетами [47]. Высокая чувствительность пейсмекеров к слабым внешним сигналам достигается за счет кооперативных эффектов в упорядоченных пучках нейронов супрахиазматических ядер, пучка Гиса и ядер ретикулярной формации.
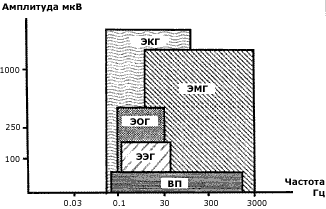
Рис.20. Амплитудно-частотные соотношения биоэлектрических сигналов [46]
Спектр частот ритмики мозга отвечает электрической активности различных структур в иерархии мозга (Рис. 20). К устойчивым ритмам спектра ЭЭГ относят: дельта-ритм (0,5-4 Гц); тэта-ритм (5-7 Гц); альфа-ритм (8-13 Гц); мю-ритм – по частотно-амплитудным характеристикам сходен с альфа-ритмом, но преобладает в передних отделах коры больших полушарий; бета-ритм (15-35 Гц) и гамма-ритм (выше 35 Гц). Мю-ритм, возможно связан с электромагнитной активностью глаз и с ритмикой сканирования ЭМ-вихрем лобных долей мозга. Причиной других ритмов могут быть электромагнитные колебания, захватывающие кору и различные структуры подкорки. В принципе, для каждого типа колебаний в спектре ЭЭГ можно выделить в мозгу емкостно-индукционные LС-структуры и смоделировать их взаимосвязь эквивалентной схемой колебательного контура, имеющего по (6) свою частоту.
Например, альфа-ритм, отвечая фоновой электрической активности коры, поддерживает на должном уровне стабильность связей неокортекса и таламуса. При удалении таламуса или отсечении его связей с корой альфа-ритм исчезает. Право-левые доли таламуса и коры полушарий можно представить разноименными обкладками двух сферических конденсаторов, а нервные связи между ними (лучистости таламуса) будут моделировать омические связи и индуктивные катушки в эквивалентных схемах контуров, работающих на частоте альфа-ритма (альфа-контур) (Рис. 21). Асимметричность индуктивных элементов альфа- контуров правого и левого полушария может лежать в основе их функциональной спецификации. Частоту колебаний в альфа-контуре, по-видимому, задают ядра-пейсмекеры ретикулярной формации, тесно связанной с таламусом. Характерное время перестроек, синхронизованных с альфа-ритмом, составляет ~100 мс.
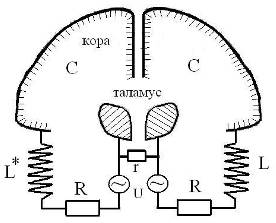
Рис .21. Эквивалентные колебательные контуры, моделирующие альфа-ритмы мозга. L, L*, R – индуктивные и омические модели лучистости таламуса (знак *) означает зеркальную инверсию хиральных структур правого полушария; r – межталамическое сращение; С и U – емкость и разность потенциалов между таламусом и корой.
Токи в нервных структурах ретикулярной формации и продолговатого мозга могут генерировать вихревые магнитные поля в структурах варолиева моста и мозжечка (Рис. 22). Внешний вид данного образования, в принципе, изоморфен в)-модели трансформированного колебательного контура (Рис. 1). Следовательно, пейсмекеры ретикулярной формации могут резонансно настраиваться на колебания стоячей ЭМ-волны геомагнитного поля (см. П. 2.4.2) и на регулярные возмущения геомагнитного поля Солнечной активностью или планетами.
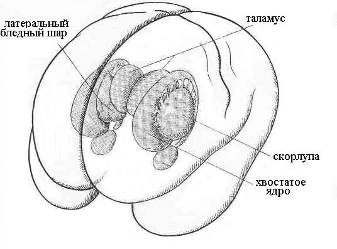
Рис. 22. Мозжечок и продолговатый мозг Рис. 23. Структуры базальных ганглий
Время усвоения одного бита зрительной информации составляет 15 – 50 мс [28]. Время элементарного мыслительного акта лежит в пределах 150 – 300 мс [48]. Из оценки скорости усвоения смысла читаемого текста, состоящего из известных слов, следует, что на осознание смысла одного слова в среднем требуется около 200 мс. Эту постоянную времени можно связать с тэта-ритмом, который манифестирует кортико-лимбические взаимодействия, регулирующие эмоции и умственную деятельность [15]. Пара эквивалентных колебательных контуров, имеющих частоту тэта-ритма (тэта-контур), будет подобна альфа-контурам (Рис.21), только взамен таламуса и его лучистости будут фигурировать соответствующие структуры гиппокампа и базальных ганглий (скорлупа, хвостатое ядро) (Рис. 23). Поскольку характерные времена передачи ПД в пределах структур мозга по порядку величины не превышают ~10 мс, то можно предположить, что скорость мыслительного акта лимитируется химическим механизмом кодирования информации, требующим активации синаптических связей.
Характерное время кинетики расходования энергоресурса мозга в процессе мышления и последующего его восстановления можно связать с кинетикой метаболизма глаз, энергетикой которых определяется интенсивность стимулирующего воздействия на лобно-височные доли мозга ЭМ-вихря (Рис. 11). Метаболизм стекловидного глаза лимитирован скоростью его гидродинамики, характерное время которой равно ~900 с. И для восстановление данного ресурса глаз достаточно дневного сна длительностью ~15 мин. Филогенетически эта постоянная времени энергетики мозга может быть обусловлена биогенным действием продуктов распада термолизованного нейтрона [6], время жизни которого равно ~900 с.
5.3. Нелокальные квантовые корреляции
5.3.1. Сознание и память
Физика сознания или самосознания, будучи тождественна физике мышления и речи, базируется на способности мозга реагировать на внешние сигналы, преобразовывать их в электромагнитные коды нейросети, запоминать и узнавать эти коды, трансформировать и синтезировать из них матрицы мыслеформ. Все эти операции возможны при наличии в мозгу физико-химических механизмов прочтения-активации и запоминания-консервации кодов. Процесс считывания и распознавания самой мыслеформы подразумевает наличие устойчивой динамичной эталонной системы кодов-реперов, составляющей физическую основу самосознания субъекта. Данная система формируется в процессе филогенеза и онтогенеза по генетической программе при непрерывном воздействии на мозг через органы чувств (зрение, слух, осязание, обоняние, вкус) всего спектра физических и социальных организующих факторов. При отсутствии последних онтогенез мозга нормального от рождения ребенка вообще не выходит на ментальный уровень. С другой стороны слепоглухонемые от рождения дети, имея только тактильный контакт с учителем, в состоянии сформировать достаточно адекватную эталонную систему кодов-реперов, позволяющую им познавать мир и общаться с людьми.
Генетическая программа кодов-реперов, определяющих индивидуальность человека, закладывается уже при формировании метрики генетического материала в процессе оплодотворения женской клетки. Развертывание данной программы на этапе эмбриогенеза подчинено закону фрактально-резонансного действия [6], который реализуется через физические механизмы энергоинформационного обмена и памяти, лежащие в основе психо- или духовно-физического изоморфизма.
Генные коды, составляя базовый уровень памяти, действуют на протяжении всей жизни человека, определяя расовые, половые и ментальные особенности его физики мозга. Следующий уровень памяти составляют безусловные рефлексы, принципы его организации закладываются на этапе морфогенеза. На этом же этапе создается химическая основа ресурса памяти первоначальных сенсорных впечатлений и ощущений. В пределах ресурса бессознательной памяти под действием социальных факторов формируется массив эталонной системы кодов-реперов сознания. Механизмы запоминания и извлечения из памяти необходимого кода-репера сочетают электрофизику и биохимию мозга с физикой динамичных систем квазифотонов, локализованных на структурно-функциональных элементах мозга.
Первый, генный уровень памяти реализуется самопроизвольно на молекулярно-клеточном уровне при синтезе ядерных ДНК, которые, в свою очередь, программируют рост и функции нервной клетки. Вообще, термодинамические особенности третичной структуры белковых молекул (ДНК, гемоглобин, ферменты и др.) позволяют им конденсировать тепловые или метаболические квазифотоны, трансформируя их в энергию колебательно-вращательных движений атомов характерных аминокислотных групп. При достижении энергии возбуждения порогового уровня она передается по фрактально-резонансному механизму на метаболит, инициируя тем самым его участие в химическом синтезе вполне определенной структуры. Так может осуществляться транслирование генной информации с молекулярного на макроскопический уровень организации живых систем.
Следующие уровни памяти образуются путем развития устойчивых нейронных связей между зонами неокортекса и базальными ганглиями, лимбическим мозгом, таламусом и мозжечком. Специализация такого блока памяти определяется функциональной доминантой второго его элемента. Учитывая синхронность морфогенеза лобно-теменных долей мозга и мозжечка, предположим, что этот блок является главным хранилищем базовых кодов-реперов сознания. Фоновая активность данного блока памяти обеспечивает непрерывность ориентации соматической и ментальной составляющих сознания в пространстве и времени. Ритмику данного режима сознания может задавать альфа-контур, обеспечивая связь между зрительной корой, мозжечком и лобными долями (Рис.24).
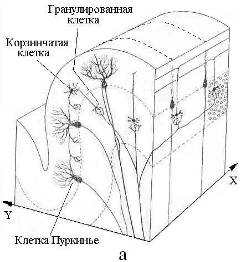
Рис. 24. Схема строения (а) и связей с корой больших полушарий (б) мозжечка – многослойного “кристалла” базового блока памяти мозга. Пунктирами показаны слои и уровни организации мозжечка
Переключение сознания на режим мышления сопряжено с заменой альфа-ритма на тэта-ритм и, соответственно, активацией блока памяти ментальных кодов-реперов, ключевыми структурами которого, очевидно, являются базальные ганглии. Можно предположить, что функциональная асимметрия мозга и сама логика мышления есть результат синергизма физики хиральных индуктивно-магнитных элементов тэта-контура (хвостатое ядро), органа обоняния (гиппокампа) и слуха (Аммонов рог) в каждом из полушарий. Участие в физике мышления емкостно-электрического элемента органа зрения (сетчатки), очевидно, заключается в активации базового блока памяти действием на лобно-височные доли мозга ЭМ-вихря глаз. Аналогичную функцию активации теменной и затылочной (предклинья, Рис. 11е) областей коры мозга, по-видимому, выполняет второй емкостно-электрический элемент зрительной системы – наружное коленчатое тело. В нем вполне может идти генерация квазифотонов-фосфенов при слиянии-рекомбинации зеркально-симметричных ЭФ от правого и левого глаза.
Физика мышления интегрирует в себе в той или иной степени функции всех структур мозга. Морфологической базой для такого объединения служит жидкостная система мозга. На начальном этапе эмбриогенеза вся нервная система локализована в нервной трубке [49], внутренний слой которой в дальнейшем превращается в односвязную внутреннюю поверхность желудочков мозга и центрального канала спинного мозга. Впоследствии ликвор, заполняя эти объемы и сообщаясь с цистернами и наружными оболочками мозга (Рис 16), берет на себя роль интегрирующей “шины”, на которую, так или иначе, замкнуты все функциональные элементы нервной системы.
5.3.2. Механизм квантовых корреляций
Следуя логике работы [50], иерархию физических состояний структурно-функциональных блоков мозга смоделируем многоуровневым фракталом из локальных динамичных систем квазифотонов. Базовой единицей данного фрактала, начиная от уровня нейросети, кончая полушариями мозга, будет LC-структура, изоморфная колебательному контуру (Рис. 8, 21). Геометрические и электрофизические параметры LC-структур будут определять тип и энергию динамических квазифотонов, а их действие формально должно удовлетворять фрактально-резонансному принципу [6]. Из цепочек LC-контуров состоят самовозбуждающиеся (реверберирующие по принципу обратной связи) замкнутые цепи в нейросетях, которые могут соответствовать образам восприятия или образам оперативной памяти. Данные возбуждения можно смоделировать затухающими колебаниями, связав их цикличность и частоту с добротностью и резонансными частотами LC-контуров, а также с длиной и степенью разветвленности петли нейросети. Для моделирования кооперативных свойств систем однотипных квазифотонов помимо электродинамики можно, в принципе, привлечь квантовую механику (Бозе-конденсация, спутанность состояний), физику волновых и нелинейных процессов (интерференция, голография, когеренция).
Предельное время психической реакции или мыслительного акта имеет порядок ~100 мс. За это время в мозгу задействуются десятки тысячи синаптических связей и активизируются десятки зон коры и структур подкорки. Даже при условии параллельности операций в нейросети и высокой скорости ассоциативных и рефлексивных связей между зонами коры и блоками памяти для обеспечения целенаправленности функциональной активности мозга в нем должен работать квантовый механизм поиска и отбора информации, кинетика которого не должна ограничиваться скоростями переноса вещества и энергии. Морфологической основой данного механизма является односвязность водной среды мозга и фрактальность его ЭМ-структуры. К примеру, каждый слой базового блока памяти (Рис. 24), состоящий из пары – древовидная клетка Пуркинье и лиановидный нейрон [49], можно считать изоморфным всему фракталу нервных связей неокортекса и позвоночника вкупе с блуждающим нервом (вагус). Изоморфизм нервная клетка – макроструктура обеспечивает высокую оперативность базового блока памяти кодов-реперов при предельно большом значении его удельной емкости.
Взаимосвязь квантовых систем может быть двух типов. Первый предполагает наличие перекрывания волновых функций электронов двух пространственно разделенных структур нервной системы мозга. В энергоинформационном обмене при этом участвуют как квазифотоны различного типа (солитоны, фононы, ЭМ-кванты), так и электрон или протон (туннельные переходы). Такая связь, по сути, аналогична механизму сальтаторной проводимости и эффективна для химически связанных систем. Второй тип связи – нелокальных квантовых корреляций [7] предполагает информационный обмен между изоморфными структурами мозга по механизму фрактально-резонансной связи [6]. Данный механизм можно распространить и на обмен информацией между мозгом и внешней средой. Действует он и на молекулярном уровне при репликации ДНК, и на социально-биологическом уровне между матерью и детьми (особенно с дочерью).
В общем случае для реализации механизма нелокальных корреляций системы должны удовлетворять двум требованиям. Первое состоит в том, что в зародышевом состоянии они должны образовывать односвязную систему, находящуюся в квантовом состоянии, определяемом тем или иным типом взаимодействия (ядерное, электромагнитное, слабое, гравитационное, биологическое, социальное). Примеры: позитроний, радиоактивное ядро, протозвезда, молекула ДНК, нервная трубка, эмбрион и организм матери. Второе требование называется несепарабельностью или запутанностью состояний химически не связанных систем и заключается в наличии механизма квантовой корреляции физических характеристик состояний. Для перечисленных выше систем их дочерними коррелирующими подсистемами будут, соответственно: два фотона, продукты радиоактивного распада, двойные звезды или звездно-планетарные системы, расщепленная ДНК, мозжечок и неокортекс, мать и дитя.
Механизм квантовой запутанности подразумевает сохранение информационной связи между дочерними подсистемами на уровне взаимодействий, определяющих собственную целостность подсистем. Универсализм квантовой запутанности генетически обусловлен подчинением процесса образования дискретных форм материи, а затем и элементарных частиц механизму бутстрапа [6]. На уровне элементарных частиц и ядер, на котором нелокальные корреляции экспериментально наблюдаются [7], природа информационных взаимодействий еще не установлена, можно только предполагать, что их кинетика лимитирована скоростью передачи импульса в физическом вакууме (эфире) ~1022 см/с [6]. С такой же скоростью, очевидно, осуществляется обмен информацией на уровне социальных и космических подсистем.
За основу механизма запутанности структурно-функциональных подсистем мозга, можно принять односвязность его водной основы, благодаря которой между системами квазифотонов возможен обмен информации со скоростью V = C/n (5). Подчинение данного обмена фрактально-резонансному принципу действия сводит вероятностный его характер к целенаправленному отбору между изоморфными структурами пары, для которой уровень запутанности состояний квазифотонов максимален.
6. ЗАКЛЮЧЕНИЕ
Дж. Максвелл полтора века назад связал развитие фундаментальной физики с познанием мозга [5]. Более 30 лет назад Ф. Крик предположил, что молекулярная биология вполне может установить биохимические основы самых сложных духовно-ментальных функций мозга, достигнув при этом уровня “биохимической теологии” или “теохимии” [51]. Однако в 2000-ом году Ф.Крик уже воздержался от прогнозирования развития биохимии мозга, подчеркнув лишь большое “значение нелинейных динамических систем в биологии на всех уровнях”. Данное уточнение вполне можно принять за адаптацию идеи Дж. Максвелла к биологическим объектам и заключить, что переход биохимии мозга в “теохимию” может осуществиться только одновременно с заменой материалистической парадигмы физики на духовную [1]. Другими словами физика мышления может состояться только как естественная теология или теофизика [52]. Эволюция естествознания уже имеет один диалектический скачок в виде перехода от классической физики к физике квантовой. Очевидно, что в недрах естествознания назрел следующий скачок – от квантовой физики к физике духа или теофизике. При всей драматичности такого скачка для физиков-идеалистов и психологов-материалистов он неизбежен. С учетом этого в настоящей работе, проанализировав физические особенности структур мозга и его кинетических характеристик, показали принципиальную возможность формализации физики мышления с помощью экстраполяций достоверных закономерностей классической и квантовой физики на субэлементарный уровень организации материи.
ЛИТЕРАТУРА
1. Холманский А.С., Стребков Д.С. Энергетика ноосферы // Доклады РАСХН. 2004. № 1.
2. Эйди У.Р. Кооперативные механизмы восприимчивости мозговой ткани к внешним и
внутренними полям. М., Физиология человека. 1975
3. Холманский А.С. Зависимость от температуры оптической активности
физиологических растворов сахаров // Математическая морфология. Электронный
математический и медико-биологический журнал. – Т. 5. – Вып. 4. - 2006. -URL:
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky/holmansky.htm
Кинетический фактор фотосинтеза // там же
4. Бехтерева Н. П., и др. Нейрофизиологические механизмы мышления. Л., 1985. 272 с.
5. Максвелл Дж.К. Избранные сочинения по теории электромагнитного поля. М. 1954
6. Холманский А.С. Энергоформа // http://www.sciteclibrary.ru/rus/catalog/pages/7441.html
Фрактально-резонансный принцип действия // http://ikar.udm.ru/mis-rt.htm
7. Менский М.Б. Концепция сознания в контексте квантовой механики //УФН, 2005.
Т.175. № 4; Доронин С.И. Мера квантовой запутанности чистых состояний // Квантовая
Магия, том 1, вып. 1. http://quantmagic.narod.ru/volumes/VOL112004/abs1123.html
8. Кобозев Н.И. Избранные труды. Т. 2. МГУ. 1978. 396 с.
9. Татур Т. А. Основы теории электромагнитного поля. М. 1989
10. Холодов Ю. А., и др. Магнитные поля биологических объектов. М.: Наука, 1987.
11. Ристо Наатанен, Внимание и функции мозга. МГУ. 1998
12. Ф.Хухо Нейрохимия, Основы и принципы. М. Мир 1990
13. Сотников О.С. Динамика структуры живого нейрона. М.: Наука, 1982.
14. Matveev V.V., Revolution and counter revolution in cell physiology. Cell Biol. Int. 2002, 26; http://bioparadigma.narod.ru/reviewonpollack.htm
15. Физиология Человека, ред. Р. Шмидт, Г. Тевс. 1- 3 т., М., 1996
16. Poliak S., Peles E. Nature Reviews Neuroscience V. 4. № 12. Р. 968-980 (2003)
17. Албертс Б. и др. Молекулярная биология клетки.
Т 3. М., Мир, 1994
18. Терехов С.А., Лекции по теории и приложениям
искусственных нейронных сетей.
Снежинск, 1994-1998.
19. Отелин А.А. Тельце Фаттер Пачини. Л. 1976
20. Katila T. et al. Magnetic fields produced by human eye // J. Appl. Phys. 1981, V. 52 (3) 2565
21. Williamson S.J., Kaufman L., Brenner D. Evoked neuromagnetic fields of the human brain // J. Appl. Phys. 1979, V. 50 (3), 2418 - 2421
22. Введенский В. Л., Ожогин В. И. Сверхчувствительная магнитометрия и биомагнетизм. М. 1986
23. Раевская О. С. Геомагнитное поле и организм человека // Успехи физиологических
наук. 1988. т. 19. № 4
24. Холодов Ю. А., Лебедева Н. Н. Реакции нервной системы человека на
электромагнитные поля. М. 1992
25. Фениш Х. Карманный атлас анатомии человека. Минск 1997
26. Хьюбел Д. Глаз, мозг, зрение, М., 1990
27. Соловьев А. Н. К вопросу о механизме биологического действия импульсного
магнитного поля // ДАН СССР, 1963, т. 149, № 2; Аlexander H. S. The biological effekts
of magnetic fields // Am. J. Med. Electronics, 1962, v. 1, № 4
28. Хацевич Т.Н., Медицинские оптические приборы: Физиологическая оптика:
Учебное пособие. - Новосибирск: СГГА. - 1998. - Ч.1. - 98
с.
29. Холманский А.С. Оптическая активность сахара
и космофизика //
http://www.kubstu.ru/fh/fams/vipusk3.htm
30. Baker R., Magnetic bone in sinus of man // Nature, 1983, v. 301, p. 78-80
31. Брагина Н. Н., Доброхотова Т. А. Функциональные асимметрии у человека, М., 1981
32. Ларри Кэхилл, Его мозг, ее мозг: http://www.nanonewsnet.com
33. Лифшиц И. М., Квазичастицы в современной физике, в сб.: В глубь атома, М., 1964
34. Кизель В.А. Практическая молекулярная спектроскопия. М. МФТИ. 1998. 254 с.
35. Кассиль Г. Мозговой барьер // Наука и жизнь. 1986. №11
36. Н.А.Темурьянц и др. Сверхнизкочастотные электромагнитные сигналы в
биологическом мире. Киев: Наукова думка, 1992.
37. Кудряшов Ю.Б. Биофизические основы действия микроволн. – М.: МГУ. 1980
38. Синицын Н.И. и др. // Биомедицинская радиоэлектроника. –1998. – №1.
39. Холманский А.С. Ядерно-химический катализ //
http://www.sciteclibrary.ru/rus/catalog/pages/6303.html
40. Мусил Я., Новакова О., Кунц К. Современная биохимия в схемах. М.Мир. 1984, 214 с.
41. Холманский А.С. Термодинамические особенности воды и биоэнергетика // Доклады РАСХН, 2006. 2. http://www.sciteclibrary.ru/rus/catalog/pages/7897.html
42. Эткин В.А. О специфике спин-спиновых взаимодействий;
Об ориентационном взаимодействии спиновых систем. НиТ, 2002.
43. Ковальзон В.М. Мелатонин – без чудес // Природа, 2004. № 2
44. Хелимский А. М. Эпифиз, М, 1969
45. Каталымов Л. Л. Физиология возбуждения и торможения, Ульяновск, 1997
46. Гнездицкий В.В. Вызванные потенциалы мозга в клинической практике. Таганрог:
ТГТУ, 1997.
47. Биологические ритмы, ред. Ю.Ашофф. 1, 2 т. М. Мир, 1984
48. Иваницкий А.М., Стрелец В.Б., Корсаков И.А. Информационные процессы мозга и
психическая деятельность. М.: Наука, 1984.
49. Коуэн У. Развитие мозга. В сб. Мозг. М. 1984
50. Peled A., Geva A.B., Brain Organization and Psychodynamics // The Journal of
Psychotherapy Practice and Research 1999; 8:24-39.
51. Крик Ф. Молекулярная биология в 2000 году // Nature. 1970. v. 228. p. 613-615.
52. Холманский А.С., Начала Теофизики. М. 1999. 126 с.
www.portalus.ru/modules/psychology/rus_readme.php?subaction=showfull&id=1132
ABSTRACT
MODELING PHYSICISTS BRAIN
Kholmanskii A.S.
The physics of the brain of the man has two forming - a base physicist general for all mammalia and physicist of the thinking, inherent only man. The development mental constituting structured-functional organization of the brain in phylogenesis have ed with chiral factor of the external ambience, and in ontogenesis - with social factor. In base of sensitivity of the brain to these factor have placed unitied its water base, mechanism to electromagnetic induction and particularities of the thermodynamics of the brain able night appear in the dreams. For the reason unifications of the description of the mechanism of the electromagnetic processes in brain entered the concept of quziphoton, uniting at itself all forms of excitation electronic and molecular-celluar structures of the brain. The equivalent schemes oscillatory sidebar element neirosite and macrostruture brain are offered. They are made estimations kinetic parameter (the energy to activations, velocities) of the physical processes, being the basis of energy and information changing the brain with external environment. The discussed mechanisms of the work operative (physical) and constant (chemical) to memories of the brain, including model nonlocal quantum correlation.
ГНУ ВНИИ электрификации сельского хозяйства, Москва
Поступила в редакцию 10.06.2006.