УДК 515.015
РЕСУРС ФУНКЦИОНАЛЬНОЙ АСИММЕТРИИ МОЗГА
© 2006 г. Холманский А. С.
Для выяснения роли электрофизики в функциональной асимметрии мозга (ФАМ) проанализировали известные данные и с помощью оригинальной методики изучили зависимость ФАМ от физического состояния человека. Мониторинг знака доминантности мозга показал зависимость ФАМ от геокосмической обстановки. Обнаружили переменную составляющую ФАМ и назвали ее ресурсом функциональной асимметрии мозга (РФАМ). Исследовали связь РФАМ с речевой функцией мозга. Соотнесли РФАМ с асимметрией электрических потенциалов полушарий мозга и положили в основу механизма генерации РФАМ асимметрию физико-химических свойств ликвора правого и левого полушарий мозга.
1. ВВЕДЕНИЕ
1.1. Асимметрия электрофизики мозга.
Между корой и структурами подкорки существует постоянная разность потенциалов порядка нескольких мВ, которая меняется с частотой меньшей частоты ЭКГ. Во сне потенциал становится положительным, а при пробуждении или увеличении поведенческой активности бодрствующего животного – отрицательным. Общепризнанного механизма возникновения этих потенциалов нет. Предполагают, что их сдвиги в отрицательную сторону обусловлены, прежде всего, деполяризацией апикальных дендритов в слоях I и II, вызванной активностью неспецифичных таламических афферентов [1].
Наличие данных переменных потенциалов позволило в работе [2] смоделировать соответствующие структуры мозга в виде емкостных элементов эквивалентных колебательных контуров. Разрядка или перезарядка этих элементов, очевидно, сопряжена с прохождение тока в структурах, которые в эквивалентных контурах играют роль катушек индуктивности. Возникающее при этом магнитное поле по закону магнитной индукции будет генерировать вихревые токи в структурах, входящих в смежные или отдаленные эквивалентные контуры, тем самым, активизируя их. По такой схеме может идти энергоинформационный обмен внутри полушарий.
Данную электрофизическую схему, в принципе, можно распространить и на межполушарные взаимодействия, учитывая, что полушария моделируются парой параллельно связанных колебательных контуров [2]. При нагрузках, связанных со специализированной деятельностью одного их полушарий, происходит изменение асимметрии межполушарных отношений [3]. Исследования локального мозгового кровотока, интенсивности потребления глюкозы мозгом и ЭЭГ обнаруживают большую активность правого полушария при решении визуально-пространственных задач, а левого полушария – при произнесении слов в уме или шепотом, а также при чтении [4]. Самый большой коэффициент асимметрии, в том числе и морфологической, отмечен в зонах Брока и Вернике, получающих максимальное развитие в онтогенезе [3].
Активизация одного полушария приводит к изменению уровня постоянного потенциала (УПП) головного мозга, который может служить независимой характеристикой интенсивности межполушарных взаимодействий [5]. Более высокий, в среднем, УПП у правшей в различных областях левого полушария означает более высокую активность и вероятно большее число работающих нервных клеток доминантного полушария. У левшей нет достоверного преобладания УПП в одном из полушарий, при тенденции к более высоким значениям УПП в правом. Механизм возникновения УПП до конца не установлен. У здоровых людей, знак межполушарной разности УПП может изменяться при различных воздействиях и, прежде всего, в стрессовом состоянии. При этом, чем выше интенсивность нагрузки, в том числе физической, тем с большей вероятностью происходит инверсия межполушарных отношений. В норме у большинства правшей в возрасте от 20 до 60 лет по данным УПП интенсивность энергетического обмена выше в левом, доминантном полушарии. С возрастом происходит постепенное сглаживание межполушарных различий за счет снижения активности левого полушария в старческом возрасте. У неграмотных людей ФАМ меньше, чем у грамотных [1].
Эти данные согласуются с предположением о том, что в основе физики мышления лежит ФАМ [2]. Причем полноценное, творческое мышление формируется у человека только при условии интегрирования именно разных функций обоих полушарий. Если слабая выраженность ФАМ характерна для умственно неразвитого человека или животного, то разрыв нервных (электрических) связей между полушариями при рассечении мозга ведет к патологическому расщеплению сознания и утрате человеком способности воображать и мыслить [6].
Таким образом, известные данные свидетельствуют, что электрофизические проявления ФАМ тесно связаны с асимметрией энергетики метаболизма, которая, в свою очередь, обусловлена асимметрией энергоинформационных отношений мозга с телом и с внешней средой [7]. Механизмы адаптации организма к внешним физическим факторам (электромагнитные поля, гравитация, радиация, температура, давление), очевидно, ответственны за формирование генетических кодов ФАМ. Генезис асимметрии физики мозга, лежащей в основе функции мышления, очевидно, связан с действием на организм универсального физического фактора, обладающего собственной хиральностью (поляризованные фотоны, нейтрино) [8].
Геокосмические условия меняются с периодами в масштабе времени от реального (день, месяц, год, век), до геологического (тысячелетия, миллионы лет). Отметим, что состояние внешних факторов, влияющих на ФАМ, как правило, не учитывается при ее исследованиях, не смотря на то, что они могут вносить систематические поправки в результаты опытов.
1.2. Асимметрия биохимии мозга
Асимметрии энергетики метаболизма и электрофизики соответствующим образом проявляются и на уровне биохимии мозга [9]. Обнаружено преимущественное содержание дофамина и более высокая активность ацетилтрансферазы в левом бледном шаре по сравнению с правым [3, 9], а также более высокая активность ацетилхолинэстеразы в коре левого полушария человека по сравнению с правым. Помимо дофамина в левом полушарии больше ГАМК, ацетилхолина, а в правом – серотонина, норадреналина [10]. Установлена асимметрия в распределении пептидов, участвующих в регуляции двигательных функций. Есть указания на большее воздействие аминазина на структуры левого полушария мозга, алкоголя на структуры правого. Имеются сведения об асимметричном распределении нейрогормонов (например, гонадотропин-рилизинг-гормона), их рецепторов, а также нейропептидов и их рецепторов. Описана асимметрия в распределении опиатных рецепторов [9].
1.3. Физическая химия ликвора
Ликвор или спинномозговая жидкость (СМЖ) является энергоинформационным посредником, обеспечивающим связь мозга с внешней средой и в том числе с кровеносной системой. Ликвор влияет на физиологическое состояние мозга и активность нервной системы. Соответствуют данной роли ликвора его физико-химические свойства и особенности его анатомии. В СМЖ могут присутствовать более 50 биологически активных веществ, как образующихся в мозгу, так и поступающих из крови. Ликвор содержит кислород, углекислый газ, электролиты, хиральные вещества: белки, молочную кислоту и сахара (90% глюкоза, 10% декстроза). Причем максимальная концентрация глюкозы наблюдается в желудочках мозга [4].
СМЖ головного и спинного мозга образует односвязную и однородную жидкостную систему, обладающую хиральными и кооперативными свойствами. Данные свойства ликвора в полной мере реализуются в желудочках и цистернах, играющих роль накопителей и преобразователей электромагнитных сигналов (квазифотонов), генерируемых как структурами мозга, так и внешними источниками земной и космической природы [2].
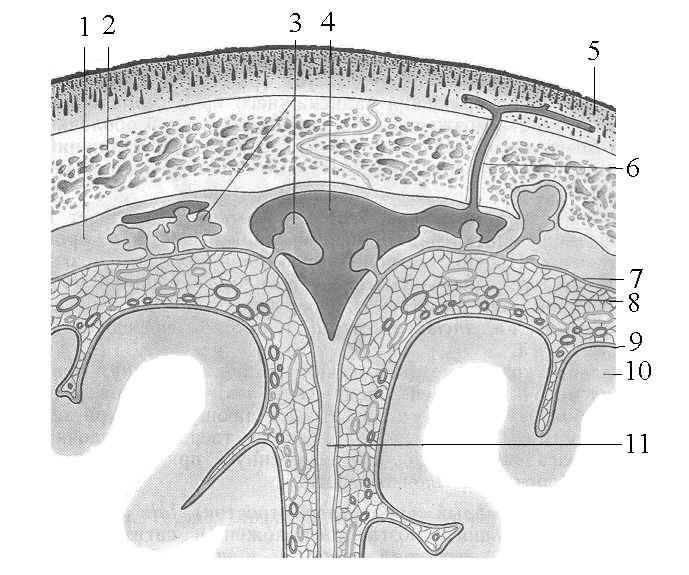
К внешним электромагнитным сигналам наиболее чувствительным должен быть ликвор, расположенный на поверхности полушарий мозга (наружный ликвор) и в центральном канале спинного мозга с его конечной цистерной. Наружный ликвор заполняет пространство между мягкой оболочкой мозга, примыкающей непосредственно к коре больших полушарий, и твердой оболочкой, связанной с костью черепа (Рис 1). Расположенная между ними проницаемая для ликвора паутинная оболочка, как трехмерная матрица играет существенную роль в термодинамике и электрофизике ликвора.
Рис 1. Схема взаимоотношений оболочек головного мозга и верхнего сагиттального синуса; разрез во фронтальной плоскости. 1 – твердая оболочка; 2 – кость черепа; 3 – арахноидальные грануляции; 4 – верхний сагиттальный синус; 5 – кожа; 6 – вена; 7 – паутинная оболочка; 8 – подпаутинное пространство; 9 – мягкая оболочка; 10 – кора больших полушарий; 11 – серп большого мозга [11].
Трехмерная вязь паутинной оболочки образована из различных соединительнотканных структур (тяжей, каналов), клеточных пятен, клеточных холмиков, ворсин и грануляций, которые могут вдаваться в субдуральное пространство и в венозные синусы (Рис 1, 2). Грануляции преимущественно расположены вдоль сагиттального синуса, поперечного синуса, у начала прямого синуса, на основании мозга, в области Сильвиевой борозды. Грануляции мягкой оболочки мозга аналогичны выростам других внутренних оболочек: ворсинам и аркадам серозных оболочек, синовиальных ворсинок суставов и другим. Оптически активные коллагенновые волокна оболочек, соединительнотканных тяжей и ликвороносных каналов могут вносить свой вклад в кооперативные свойства ликвора благодаря эпитаксиальному эффекту [2]. Кроме того, они могут играть роль хиральных фильтров-преобразователей (поляроидов) для внешнего МКВ-излучения [13], поглощаемого ликвором и затем верхними слоями коры мозга.
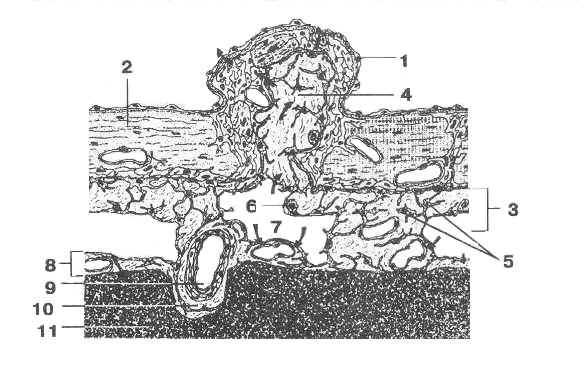
Рис 2. Мозговые оболочки. 1 – эндотелиальная клетка; 2 – твердая мозговая оболочка; 3 – паутинная оболочка; 4 – соединительная ткань; 5 – клетки паутинной оболочки; 6 – макрофаг; 7 – подпаутинное пространство; 8 – мягкая мозговая оболочка; 9 – кровеносный сосуд; 10 – перивапскулярная соединительная ткань; 11 – мозг [12].
Особого внимания заслуживают грануляции, которые развиваются у человека к 10 годам жизни. Несостоятельно с точки зрения механики отнесение их к “барорецепторам”, “вентилям” и “фиксаторам” мозга в черепной коробке [4]. Поскольку эти образования развиваются в онтогенезе синхронно с установлением альфа-ритма мозга и половым созреванием, разумно их отнести к чувствительным органам адаптационного аппарата организма, обеспечивающим энергоинформационную связь ликвора с внешней средой и кровью. Роль носителя квантов энергии (квазифотонов) между кровью и ликвором может исполнять углекислый газ или бикарбонат (НСО3–), молекулы которых в ликворе и венозной крови резонансно поглощают внешнее МКВ-излучение [2, 7]. Концентрация углекислого газа в ликворе больше чем в венозной крови, а тем более в артериальной, поэтому насыщенные энергией колебательных квазифотонов молекулы СО2 и НСО3– будут диффундировать из ликвора в кровь.
Из результатов исследования оптической активности физрастворов, содержащих глюкозу, декстран и коллагеновые волокна желатина [8], можно заключить, что ликвор, в принципе, должен обладать оптической активностью и ее знак и величина должны зависеть от температуры, хиральных метаболитов и геокосмической обстановки. В частности, понижение температуры мозга на ~1о в состоянии ночного сна, а равно, и кванты МКВ-излучения соответствующей энергии [8] вполне могут инициировать в ликворе образование хиральных надмолекулярных структур из кластеров воды и ассоциатов из сахаров. В результате этих низкоэнергетических перестроек структуры ликвора, аналогичных фазовым переходам в жидких кристаллах и гелях, может возрастать степень упорядоченности ликвора и меняться его электрофизические свойства.
ФАМ в определенной степени является производной асимметрии химических и физических свойств ликвора подпаутинных пространств правого и левого полушарий, бассейны которых практически не смешиваются. Очевидно, вносит свою лепту в ФАМ и асимметрия свойств ликвора не сообщающихся напрямую боковых желудочков мозга. Установлено [4] различие химического состава ликвора разных полушарий, а также наличие в нем носителей патогенной информации, избирательно действующих на нейроны правой и левой половин тела. Поскольку процессы самоорганизации хиральных растворов чувствительны к добавкам органических веществ и электролитов [8], можно предположить, что и кооперативные свойства ликвора в правом и левом полушарии будут различаться. Данные различия должны усиливаться во время ночного сна, вследствие снижения температуры коры мозга и изменения режима метаболизма мозга [14].
С целью выяснения роли электрофизики в механизмах обработки и передачи информации в мозгу в настоящей работе исследовали характер зависимости ФАМ от различных внешних и внутренних факторов.
2. ЭКСПЕРИМЕНТ
Зависимость функционального состояния мозга от внешних условий изучали с помощью метода [15]. В его основе лежит измерение скорости и направления вращения человека совершающего бег на месте либо на неподвижном основании с закрытыми глазами (абсолютный метод), либо на подвижном коврике с открытыми глазами (относительный метод). Относительный метод использовали при кинетических исследованиях, определяя скорость вращения (W) по величине угла поворота коврика за время бега в течении 1 – 3 минуты. При этом ориентация человека лицом на восток оставалась неизменной. В опытах участвовали мужчина (~50 лет, правша) и мальчик (~7 лет, правша).
Известно [3], что в норме у правши речевой центр расположен в левом полушарии, при этом правая рука и в меньшей степени правая нога оказываются ведущими. При соблюдении этих условий левое полушарие называют доминантным. Отсюда следует, что при совершении бега на месте правша в норме должен вращаться против часовой стрелки. Будем считать угловую скорость (W) такого вращения положительной. Вращение же его по часовой стрелке (W – отрицательна) свидетельствует об инверсии межполушарных отношений (П. 1. 3). Формально, оставляя вопрос о локализации речевой функции открытым, правое полушарие в этом случае следует считать доминантным.
На Рис 3 приведены результаты мониторинга направления вращения правши на протяжении 10 лет. Смена направления вращения, а значит, и активного полушария, по-видимому, была синхронизована с изменением расположения Земли, Солнца и планет. Например, обращение знака скорости вращения 20.06.06 и последующее ее зануление совпало с летним солнцеворотом 21.06.06. Абсолютная величина W также менялась со временем в диапазоне от 0 до ~? рад/мин, что может быть обусловлено изменением интенсивности действия физического фактора на человека. Следует отметить, что по абсолютной величине W в годы, предшествующие последнему максимуму солнечной активности (1997 – 2000 гг.), была существенно больше, чем в годы спада солнечной активности (2001 – 2006 гг.). Более того, в процессе бега на улице с чтением стихов про себя в определенные дни до миллениума наблюдали асимметричное и стойкое покраснение кожи в области темени, что свидетельствует о зависимости гемодинамики в районе боковых лакун сагиттального синуса от геокосмической обстановки.
Эти результаты согласуются с предположением о зависимости ФАМ от гелиофизического фактора [16]. На его вариации в реальном масштабе времени должна соответствующим образом реагировать асимметрия активности мозга вплоть до ее инверсии. Например, в работах датированных 1977 – 1983 гг. наблюдали преобладание активности правого полушария во время сновидений и быстрого сна [17]. Однако результаты исследований 1984 – 1990 годов показали отсутствие функциональной асимметрии полушарий во время быстрого сна, при этом межполушарная когерентность ЭЭГ на всех стадиях сна была выше, чем в состоянии бодрствования [17]. Такая невоспроизводимость результатов может быть обусловлена инверсией межполушарных отношений в указанные периоды времени.
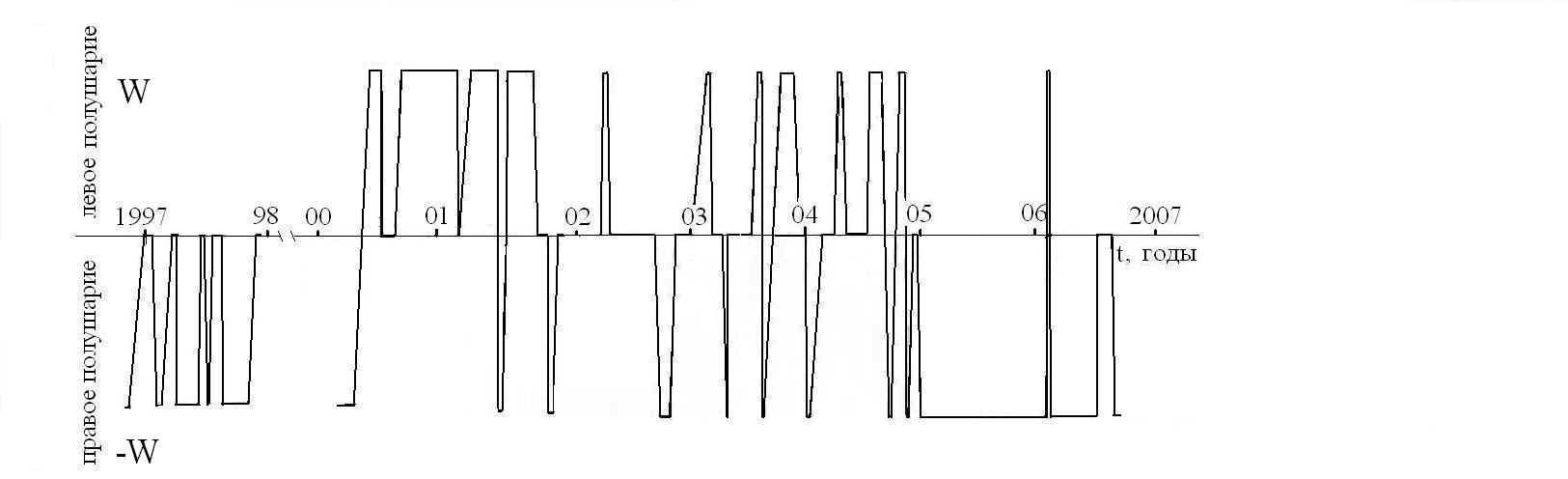
Рис 3. Зависимость знака доминантности мозга мужчины (правши) от времени.
Исследования показали, что для количественных оценок характера и кинетики изменений ФАМ можно использовать величину скорости вращения (W) человека совершающего бег на месте. Как правило, величина W имела некое максимальное значение утром и в начале опытов, а затем снижалось со скоростью зависящей от различных условий. С максимальной величиной W, фиксируемой в начале опыта, соотнесли ресурс функциональной асимметрии мозга (РФАМ), полагая, что его электрофизическая составляющая равнозначна УПП. Знак РФАМ менялся синхронно с изменением доминантности мозга (Рис 3), а его максимальное значение, как правило, наблюдалось сразу после ночного сна или после дневного сна-дремы. На это указывает монотонное снижение скорости вращения при повторных опытах (Рис 4 – 6). К расходованию РФАМ ведет физическая нагрузка (Рис 4, 5), тогда как чтение наизусть стихов (Евангелие от Матфея глава 6, стихи 9 -13) в процессе бега активизировало РФАМ (Рис 7) и замедляло кинетику его снижении (Рис 5).
Кинетика снижения РФАМ (W) качественно коррелирует с кинетикой изменения угла вращения оптически активного физиологического раствора глюкозы (Рис 6), что позволяет предполагать участие в накоплении РФАМ жидких сред организма, содержащих сахара.
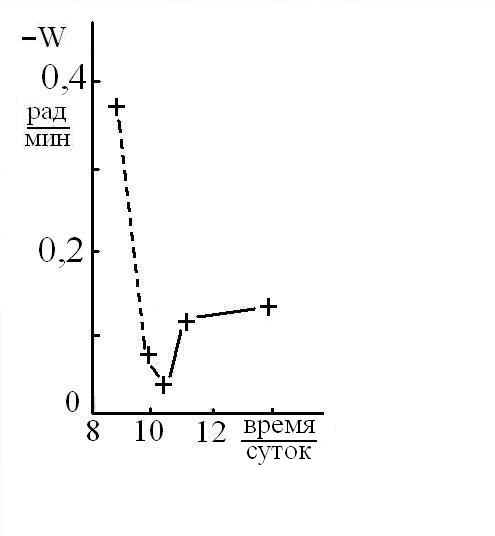
Рис 4. Кинетика спада в течение дня (16.03.05) скорости вращения мужчины по часовой стрелке, совершающего бег на коврике и читающего про себя стихи. Пунктиром обозначен 30-минутный бег на улице.
Качественные оценки влияния на кинетику спада РФАМ (W) продуктов питания, спиртных напитков и дневного сна-дремы показали, что потребление сахара, винограда, виноградного вина (каберне, кагор – 50 гр) в процессе опыта ведет к увеличению W. Такой же эффект дает и дневной сон-дрема длительностью 15 мин, тогда как потребление водки или коньяка (50 гр) приводило к снижению W. Эти результаты согласуются с известными данными об асимметрии действия лекарств и алкоголя на полушария (см. П 1.2), с учетом которых можно заключить, что повышенное действие этанола водки и коньяка на правое полушарие ведет к снижению W.
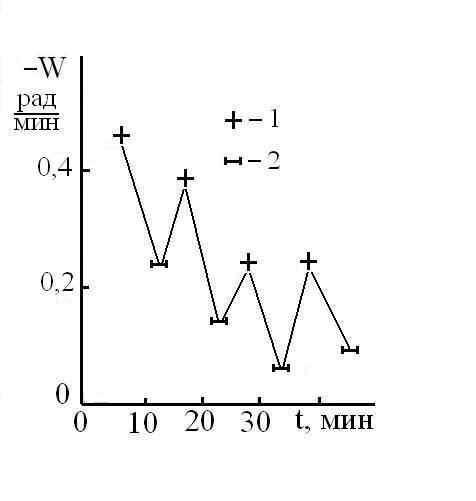
Рис 5. Изменение скорости вращения по часовой стрелке мужчины, совершающего бег на коврике с чтением про себя стихов (1) и при рассеянном состоянии мозга (2). Первая точка – 10 часов 12.03.05.
Очевидно, что органические биогенные компоненты виноградных вин и сахар в большей степени активизируют левое полушарие (см. П 1.2), что и приводит к увеличению W. По той же причине W может возрастать при чтении стихов (П 1.1; Рис 5).
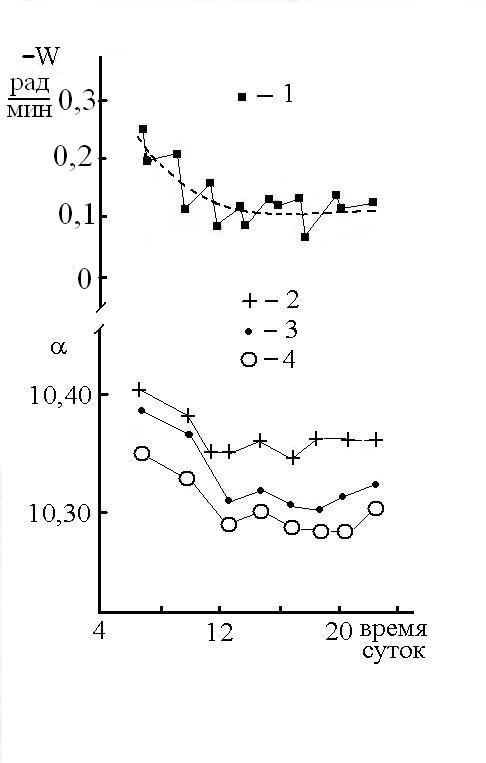
Рис 6. Изменение скорости вращения мужчины, совершающего бег на коврике и читающего про себя стихи (1); пунктир – средняя линия. Зависимость величины угла вращения физраствора глюкозы (10%) от времени суток (14.03.05). Поляриметр СМ-3 ориентирован по вертикали к поверхности земли (2), на восток (3) и на север (4).
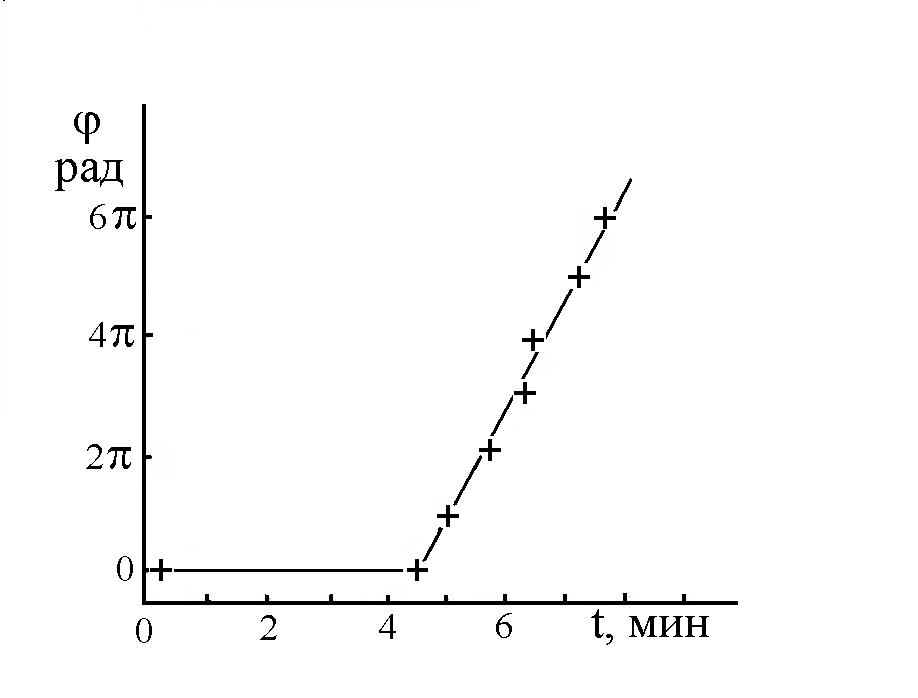
Рис 7. Зависимость величины угла вращения по часовой стрелке мальчика семилетнего возраста, совершающего бег на неподвижной основе. Нулевая точка 10 часов 05.12.97. От 0 до 4,5 мин бег без чтения стихов, от 4,5 мин – бег с чтением стихов.
Для оценки асимметрии энергетики метаболизма на уровне сомы измерили скорость роста ногтей на больших пальцах ног и рук мужчины (Рис 8). Исходя из полученных результатов, можно полагать, что незначительная асимметрия клеточного метаболизма наблюдается только на уровне рук и практически отсутствует на уровне ног. Данный результат согласуется, по крайней мере, с распределением капсулированных нервных окончаний (тельца Пачини), которых в правой руке правши заметно больше, чем в левой, тогда как на подошвах ног такой разницы не наблюдается [18]. Кроме того, следует отметить, что сравнивание скорости роста ногтей на правой и левой руке после июня могло быть обусловлено изменением гелиофизического фактора 22.06.06. Можно заключить, что вариации РФАМ, регистрируемые по W, не связаны с асимметрией клеточного метаболизма в мышцах ног и обусловлены, очевидно, асимметрией энергетики нервной системы организма, производной от ФАМ.
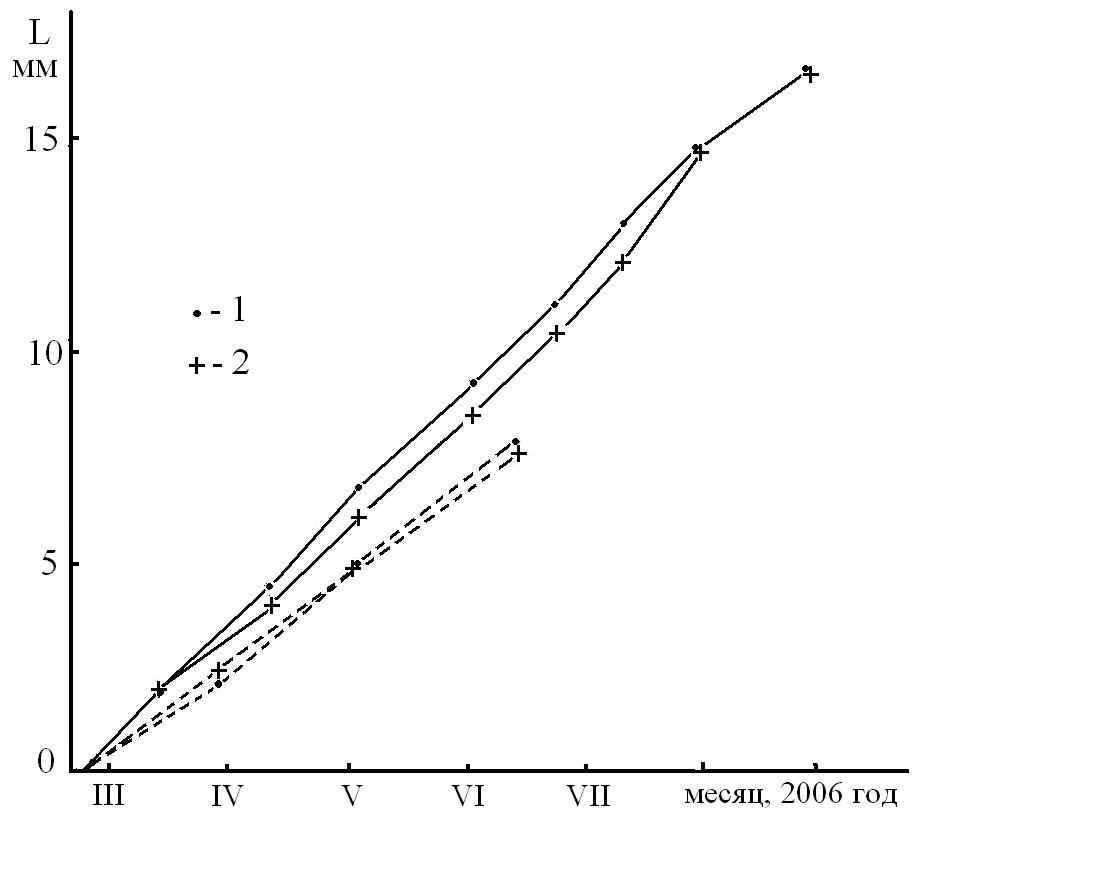
Рис 8. Скорость роста ногтей на больших пальцах рук (сплошные линии) и ног (пунктирные линии) правши. 1 – правые пальцы; 2 – левые.
3. ОБСУЖДЕНИЕ
Обнаруженный нами РФАМ в совокупности с известными данными об электрических потенциалах и магнитных полях мозга (см. П. 1.1 и ссылки в [2]) вполне могут быть использованы для обоснования предложенных в [2] физических моделей механизма передачи информации между структурами мозга. Об этом свидетельствует установленная нами связь РФАМ с речевой функцией мозга. Отметим, что в опытах читали про себя, как правило, стихи из Евангелия от Матфея, которые известны как молитва “Отче наш”. Автоматизм чтения наизусть, обеспеченный хранением стихов в оперативной памяти и многолетней практикой их повторения, очевидно, равнозначен спонтанной речи.
Двигательная область коры больших полушарий (поле 6 на карте коры [10]), отвечающая, в частности, за повороты тела, входит в состав лобных долей мозга, но расположена вне радиуса действия вихревого магнитного поля глаз [2]. Стимуляция электрическими импульсами нейронов поля 6 вызывает вращение туловища, глаз и поднятие контралатеральной руки [10]. Причем повреждения, затрагивающие двигательную область коры, нарушают и спонтанную речь [10]. Отметим, что над полем 6 расположен бассейн ликвора (рис 9) и боковые лакуны сагиттального синуса (Рис 1). Учитывая эти анатомические данные и полученные в работе результаты, можно построить электрофизическую модель генерации РФАМ (УПП), положив в ее основу асимметрию физико-химических свойств ликвора в правом и левом полушариях.
Асимметрия кооперативных свойств ликвора подпаутинного пространства (бассейны на Рис 9) должна сказаться на эффективности процессов образования надмолекулярных структур в условиях ночного сна (П 1.3). Соответственно, будет различаться и степень поляризации наружного ликвора в полушариях, а значит, и распределения зарядов на мягкой оболочке со стороны ликвора. Индуктивно это различие в распределении зарядов отобразится и на другой стороне мягкой оболочки, то есть на поверхности I-го слоя коры, включающего нейроны ориентированные параллельно поверхности мягкой оболочки (П 1.1). Дифференциация распределения зарядов на поверхности коры полушарий обусловит различие УПП внутри и между полушариями и, соответственно, асимметрию в активности нейронов поля 6.
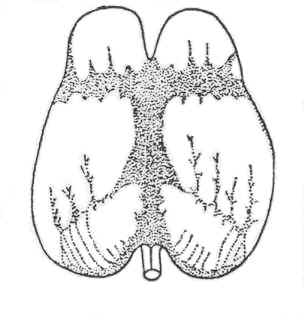
Рис 9. Ликворные бассейны на наружной поверхности головного мозга [19].
Известно [14], что углекислый газ является эффективным регулятором функции дыхания и кровоснабжения мозга. Приток крови к мозгу существенно возрастает при повышении содержания СО2 в крови. При этом интенсифицируется метаболизм мозга и возрастает концентрация ионов Н+ и К+ [14]. Динамика диффузии СО2 из ликвора в кровь через грануляции (П 1. 3) в полушариях вполне может оказаться зависима от внешних и внутренних факторов асимметрии (П 1.3). В таком случае возникает дополнительный фактор асимметрии метаболизма мозга, который приведет к асимметрии распределения ионов в полушариях, а значит, усилит УПП и РФАМ.
Как отмечалось выше (П 1.3) зависимость физико-химических свойств наружного ликвора от внешних физических факторов максимальна в ночное время в состоянии сна. Вариации доминантности мозга (Рис 3) и утренней величины W говорят о том, что данные факторы обладают переменной хиральностью и интенсивность их действия на правое и левое полушария определяется конкретной геокосмической обстановкой.
Увеличение РФАМ в процессе бега у взрослого и его активация у ребенка чтением стихов, очевидно, обусловлено передачей по механизму магнитной индукции возбуждения от центров спонтанной речи на центры двигательной коры, отвечающие за вращение. При этом факт усиления (у взрослого) и включения (у ребенка) скорости вращения по часовой стрелке при чтении стихов указывает на такое усиление асимметрии мозга, при котором возрастает активность именно правого полушария. Поскольку чтение про себя активизирует левое полушарие (П 1.1.), то надо полагать, что при этом снижается активность центров вращения данного полушария и функция правого полушария начинает доминировать.
4. ЗАКЛЮЧЕНИЕ
Полученные в настоящей работе результаты, согласуясь с известными данными о функциональной асимметрии мозга, углубляют понимание электрофизики мозга, ответственной за энергоинформационные связи как внутри мозга, так и между мозгом и внешней средой. Это говорит о перспективности разработанного нами метода исследования ФАМ. Выявленный в работе ресурс функциональной асимметрии мозга и его связь с речевой функцией мозга позволяют положить в основу физики мышления межполушарную функциональную асимметрию. Установленная в работе зависимость ресурса ФАМ от внутренних и внешних условий позволяет высказать гипотезу о том, что эволюционным итогом периодической инверсии асимметрии мозга явится усвоение правым полушарием функции речевого аппарата, что будет гарантировать человеку сохранение его мозгом способности плодотворно мыслить при любой геокосмической обстановке.
ЛИТЕРАТУРА
1. Фокин В.Ф. Пономарева Н.В., Городенский Н.Г. и др. Функциональная
межполушарная асимметрия и асимметрия межполушарных отношений.
“Системный подход в физиологии”. – 2004, №12.- С. 111-127.
2. Холманский А.С. Моделирование физики мозга // Математическая морфология.
Электронный математический и медико-биологический журнал. Т. 5. В. 4. 2006.
http://www.smolensk.ru/user/sgma/MMORPH/N-12 html/holmansky/holmansky.htm
3. Брагина Н. Н., Доброхотова Т. А.Функциональные асимметрии человека. – М.:
Медицина, 1981. – 288 с.
4. Ахметсафин А.Н., Ликвор // www.hanbalik.narod.ru/manual_medicine/liquor.htm
5. Пономарева Н.В., Фокин В.Ф. Уровень постоянных потенциалов мозга как
показатель интенсивности церебрального энергетического обмена при чтении
// Новое в изучении пластичности мозга. РАМН. Материалы конференции. Отделение медикобиологических наук, НИИ мозга РАМН.- М 2000.- с. 70;
Фокин В.Ф., Пономарева Н.В. Энергетическая физиология мозга, М.: 2003. 287 с.
6. Трефферт Д., Кристинсен Д. Феноменальный мозг // В мире науки. – 3. 2006
7. Холманский А.С. Термодинамические особенности воды и биоэнергетика // Доклады РАСХН, 2006. 2. // http://www.sciteclibrary.ru/rus/catalog/pages/7897.html
Зачем люди пьют и курят // http://www.sciteclibrary.ru/rus/catalog/pages/9076.html
8. Холманский А.С. Зависимость от температуры оптической активности
физиологических растворов сахаров // Математическая морфология. Электронный математический и медико-биологический журнал. – Т. 5. – Вып. 4. - 2006. -URL: http://www.smolensk.ru/user/sgma/MMORPH/N-12; Оптическая активность сахара и космофизика // http://www.kubstu.ru/fh/fams/vipusk3.htm;
9. Вартанян Г.А., Клементьев Б.И. Химическая симметрия и асимметрия головного мозга. – М.: Медицина, 1991. – 190 с.
10. Физиология человека. Ред. Р. Шмидт, Г. Тевс, Т. 1-4, - М.: Мир, 1996
11. Анатомия человека. Ред. М.Р.Сапин. Т 2. – М.: Медицина, 1993. – 560 с.
12. Гистология, ред. Э.Г. Улумбеков, – М.: Медицина. – 1998, 950 с.
13. Азанов С.В., Вольхин И.Л., Коротаев Н.Н. Вращение плоскости поляризации СВЧ волны хиральным композитом в свободном пространстве //
www.bti.secna.ru/institute_/konf/44.doc
14. Гречин В.Б., Кропотов Ю.Д. Медленные неэлектрические ритмы головного мозга человека. – Л.: Наука, 1979
15. Холманский А.С. Способ определения функционального состояния человека //
Патент РФ 2193859 от 10.07.2001. Бюл. 10.12.2002, № 34. http://www.sciteclibrary.ru/rus/catalog/pages/7046.html
16. Холманский А.С. Физические факторы асимметрии мышления //
http://filosof.net/disput/holmansky/ffam.htm; Метрика приземного эфира //
http://www.sciteclibrary.ru/rus/catalog/pages/7876.html
17. Красноперов О.В. Клиническая и психофизиологическая характеристики
диссомний у больных неврозами, Канд. Дисс. Томск – 1993
18. Отелин А.А. Тельце Фаттер-Пачини. – Л.: 1976
19. Фридман А. П. Основы ликворологии. – Л.: Медицина, 1971
RESOURCE to FUNCTIONAL ASYMMETRY of the BRAIN
Holmanskii A.S.
The electro physic dug For clarification in functional asymmetry of the brain (FAM) have analyzed the known data and by means of original methods have studied the dependency FAM from physical condition of the person. Monitoring the sign dominating brain has shown the dependency FAM from geocosmical of the situation. Have Found variable forming FAM and have named its resource to functional asymmetry of the brain (RFAM). Researched the relationship RFAM with speech function of the brain. Have Correlated RFAM with asymmetry electric potential hemisphere of the brain and have assumed as a basis mechanism to generations RFAM asymmetry physical-chemical characteristic liquor right and left hemisphere of the brain.
ГНУ ВНИИ электрификации сельского хозяйства, Москва
Холманский Александр Сергеевич (asholman@mtu-net.ru)
Поступила в редакцию 20.09.2006.