
УДК 530.1+ 539.2
ЗАВИСИМОСТЬ ОТ ТЕМПЕРАТУРЫ ОПТИЧЕСКОЙ АКТИВНОСТИ ФИЗИОЛОГИЧЕСКИХ РАСТВОРОВ САХАРОВ
© 2006 г. Холманский А. С.
Исследована зависимость от температуры оптической активности водных растворов метаболических сахаров и определена энергия активации реакции их ассоциирования в надмолекулярные хиральные структуры. Ее величина оказалась близка к значениям энергии вращательного движения и биогенного МКВ-излучения. Исходя из этого и учитывая зависимости оптической активности сахаров от времени и даты, места и ориентации прибора, приняли за основу механизма действия хирального фактора фило- и онтогенеза метаболический эффект реакции образования хиральных водно-сахарных ассоциатов.
Оптически активные D-сахара являются основным источником энергии метаболизма живых систем. Обязательное посредничество ахиральной молекулы аденозинтрифосфата (АТФ) в процессе метаболизации энергии химических связей сахаров нивелирует их хиральность, унифицируя тем самым энергию макроэргических связей АТФ. Однако роль сахаров в возникновении и развитии жизни не ограничилась их вкладом в биоэнергетику. В ходе эволюции усложнялись физико-химические свойства сахаров и, соответственно, возрастала их роль в энергоинформационном обмене живых систем с внешней средой. Моно- и полисахариды вошли в состав структурных элементов биомолекул, клеток, жидких сред, информационных систем, они регулируют иммунитет высших организмов [1]. Учитывая морфо-функциональную универсальность D-сахаров, можно предположить, что именно их физико-химические свойства обеспечивают высокую чувствительность механизма адаптации живых систем к внешнему постоянно действующему хиральному фактору (ХФ) [2]. Следует отметить, что вопрос о физической природе ХФ и механизме его влияния на биогенез до сих пор остается открытым [3]. Впервые данную проблему сформулировал Л. Пастер в XIX веке, предположив, что дисимметрия живых систем возникла на ранних стадиях биологической эволюции под действием электрических и магнитных полей космического происхождения [4]. В обоснование этой идеи в работе [4] приведен возможный механизм асимметричного действия электромагнитного поля на макромолекулу белка. Согласуется с нею и гипотеза о ключевой роли сахаров еще на пребиотическом этапе биогенеза [5]. Привлекательна также идея о нейтринной природе ХФ [6], которую можно соотнести с гипотезой В.И. Вернадского о детерминировании дисимметрии живых систем хиральностью физического вакуума (эфира) [4]. Реликтовое нейтрино, составляя энергетическую основу вакуума, вполне может обеспечивать его гравитационную и электромагнитную динамику (эффект Фарадея, например) [7].
В общем случае в основе механизма чувствительности живой системы к ХФ помимо хиральности ее элементов должно лежать то или иное взаимодействие между ними, обеспечивающее переход системы в коррелированное состояние с новым качеством. Переход в данное состояние (фазу) ведет к возрастанию порядка системы или снижению энтропии и при этом увеличивается ее устойчивость (время жизни). За перестройку порядка системы на микро и макро уровне ответственны электромагнитные взаимодействия между элементами. Динамику, кинетику, радиус действия и стереометрию упорядочивающих сил в живых системах лимитируют молекулярно-кооперативные свойства воды [8, 9]. С целью получения дополнительной информации о вкладе сахаров в механизм чувствительности живых систем к ХФ в настоящей работе исследовали температурные и концентрационные зависимости оптической активности водных и водно-этанольных растворов D-глюкозы и сахаров. Данные растворы моделировали состав физиологических жидкостей (межклеточная жидкость, лимфа, ликвор, синовия). При этом варьировали место, дату и время измерения, а также ориентацию прибора относительно частей света.
Методика
Использовали поляриметр круговой СМ-З с натриевой лампой (точность измерения 0,01о, длина волны D-линии натрия – 589 нм) и кюветы длиной 200 и 100 мм. Измерения угла вращения (?) плоскости поляризации света проводили по методике, описанной в [2]. Кюветное отделение поляриметра обматывали двумя слоями резинового шланга диаметром 10/6 мм. Для варьирования температуры использовали термостат U 7. Температуру раствора контролировали с помощью термопары медь-константан, один спай которой приклеивали скотчем к наружной стенки кюветы, а второй помещался в термос со льдом (0оС). Температуру меняли в диапазоне 7–40оС. Исследовали стандартные (изотонические) физиологические растворы (ФР) D-глюкозы (10, 40%) (удельное вращение [?]D = 52,7о [10]) и полисахарида реополиглюкина (декстран, 10%). Для сравнения использовали также масляный раствор прогестерона (2,5%), разбавленный хроматографически чистым октаном до 2%. Исследовали оптическую активность композиции: ФР + хондроитинсульфат (10%) + гиалуроновая кислота (0,8%) + сахар (10%) {ФР + ХС(10%) + ГК (0,8%) + сахар (10%)}, которая моделировала высокомолекулярное соединение, играющее активную роль в энергетике соединительной ткани [11]. Величина ? раствора ФР + ХС (10%) при комнатной температуре (Тком) составила –2,70о (кювета 100 мм). Физико-химические свойства ГК описаны в [2]. Физраствор и метаболиты брали в аптеке, этанол был марки “медицинский”. Пищевой сахар ([a]D = 66,5о [10]) растворяли в физрастворе, а также в кипяченой и отстоянной в течение суток водопроводной воде. Концентрации растворов подбирали из условия сохранения достаточной прозрачности жидкостей при максимальном значении ?. Измерения проводили на первом и девятом этаже панельного дома в комнатах, расположенных на восточной (О) и на западной (W) сторонах дома.
Результаты и обсуждение
Графики зависимости угла вращения растворов от времени и температуры приведены на Рис. 1–5, а время измерения, диапазоны температур и энергии активации представлены в Таблице 1. Отметим, что Е по порядку величины близка к энергии активации, полученной из температурных зависимостей поверхностного натяжения воды [8]. Известно [2], что результаты измерений оптической активности сахара зависят от места и времени измерения, а также от ориентации прибора. Для определения диапазона и характера этих изменений провели ряд измерений при Тком (Рис 1, 2, 5, 6). Полученные результаты, согласуясь с известными данными [2], свидетельствуют о чувствительности оптической активности растворов к положению Солнца и, по-видимому, к фазе Луны (Рис 2).
Таблица 1
УСЛОВИЯ ИЗМЕРЕНИЙ И ВЕЛИЧИНЫ ЭНЕРГИЙ АКТИВАЦИИ
N п/п |
Раствор |
Дата (2005 г) (время измерения) |
Диапазон температуры (К) |
Е (кДж/моль) |
1. |
ФР + глюкоза (10%) | 24.08 (21-22) |
290 > 312 |
0 |
2. |
ФР + глюкоза (40%) |
28.09 (23–24)
|
299 > 290 290 > 296 296 > 311 |
0,45 0,45 0,10 |
30.09 (12–13) |
294 > 288 288 > 297 297> 307 |
0,35 0,35 0,10 |
||
3. |
ФР + этанол (43%) + глюкоза (23%) | 05.09 (20–22) |
297 > 285 286 > 305 |
0,55 0,55 |
4. |
ФР + сахар (45%) |
21.09 (17 -19) |
293 > 287 289 > 316 |
0,45 0,35 |
24.09 (14–15) |
297 > 290 290 > 311 |
0,45 0,40 |
||
5. |
Вода + сахар |
8.10 (16 – 19) |
296 > 288 288 > 331 |
0,30 0,30 |
6. |
ФР +декстран (10%) |
27.08 (17 -18) |
284 > 310 |
0,30 |
14.09 (22–23) |
298 > 287 287 > 301 |
0,40 0,25 |
||
7. |
ФР + этанол (36%) + декстран (6,4%) |
25.09 (22–23) |
298 > 288 289 > 298 |
0,60 0,60 |
О чувствительности прибора к локальной анизотропии внешнего хирального фактора говорит зависимость ? для всех растворов от вариации следующих условий измерений:
1) изменение ориентации прибора – восток (О), север (N), перпендикулярно поверхности земли (Z) (Рис 1);
2) изменение места положения прибора (комнаты на О- и W-сторонах дома, Рис 5);
3) изменение высоты положения прибора относительно земли (время измерения 21- 2130 , 26.10.05, Таблица 2).
Таблица 2
ЗАВИСИМОСТЬ УГЛА ВРАЩЕНИЯ ОТ ВЫСОТЫ
Раствор |
Направление | 1 этаж | 9 этаж |
ФР + глюкоза (40), кювета 200 мм. |
Восток (О) |
43,09 |
43,07 |
Север (N) |
43,07 |
43,01 |
|
ФР + декстран (10%), кювета 100 мм |
Восток (О) | 19,30 |
19,20 |
| Север (N) | 19,21 |
19,20 |
Отметим, что все три вида зависимостей показаний прибора качественно коррелирует с аналогичными зависимостями физического состояния человека, которое контролировали по величине и направлению скорости спонтанного вращения человека вокруг своей оси, при совершении им бега на месте [12]. В работе [13] исследован дрейф нулевой точки прибора СМ-1 во времени и выявлены ритмы длительностью в сутки, неделя, месяц. Учитывая эти данные и результаты [2], провели сравнительные измерения нулевой точки прибора СМ-3, а также оптической активности глюкозы (10%) и прогестерона при Тком в двух направлениях (О, N) (Рис 1). Эти измерения показали симбатное изменение со временем величины
D
a = aN – aО для обоих растворов и для нулевой точки. Величины a и D a меняются в пределах от 0 до ~0,3% от a. Причем диапазон изменений Da раствора глюкозы в 1,5 – 2 раза превышает значения Da для прогестерона и нулевой точки, которые близки. В оптической системе прибора СМ-3 использована оптически активная кварцевая пластинка, толщиной ~0,2 мм. Оптическая активность кварца почти в сто раз выше активности сахара [14]. Вследствие этого величины D a для нулевой точки и для растворов сахаров получаются одного порядка. В основе механизма возникновения и инверсии D a может лежать чувствительность показателя преломления кварца и растворов к изменениям локальных электромагнитных параметров среды, которая в случае водного раствора явно выше, чем в случае масляного раствора или кристалла кварца.
Рис 1. Зависимость угла вращения растворов от времени и ориентации прибора (Тком).
0 – дрейф нулевой точки прибора; 1 – ФР + глюкоза (10%), (кювета 200 мм);
2 – прогестерон (2%) в смеси масло + октан (кювета 100 мм).
Ориентация прибора: точки – на восток, кружки – на север, крестики – вертикально.
Поскольку изменения a при варьировании температуры не превышали 1,5% от a, то ошибка в оценке энергии активации по температурным зависимостям a могла достигать 20%. С учетом этого для обсчета выбирали наиболее гладкие температурные зависимости. Об отсутствии заметного влияния температуры на прибор говорит нулевой наклон температурной зависимости a для физраствора глюкозы (10%) (Рис 3). Этот же результат указывает на наличие порогового значения концентрации в зависимости a от температуры и позволяет исключить эффекты мутаротации глюкозы. Оптическая активность полисахарида декстрана определяется хиральностью моносахарида мальтозы, концентрация которой в линейных полимерных цепях была эквивалентна ~40% концентрации раствора сахара. Отсутствие отклонений от прямой линии концентрационной зависимости удельного вращения во всем диапазоне концентраций сахаров позволяет исключить эффекты рассеяния света для всех растворов.

Рис 2. Зависимость угла вращения раствора ФР + декстран (10%) от времени. Измерения 02.XI.05 в кювете 100 мм. Точки – направление на восток, кружки – на север.
Стрелками показано время восхода и заката Солнца и Луны (две фазы – 0 и 1/4).
Для количественной обработки результатов предположили, что в растворах сахаров при достижении порогового значения концентрации, наряду с оптической активностью отдельных молекул свой вклад в хиральность раствора вносят и надмолекулярные их образования.
В пределе межмолекулярных взаимодействий характерных для кристалла сахара его удельное вращение приобретает анизотропию (1,6 – 5,4 град/мм для различных направлений) и почти на порядок превышает удельное вращение растворов сахара [14]. О возможности вклада межмолекулярных взаимодействий в хиральность раствора говорит еще такой факт. Высокое значение удельного вращения кристалла кварца (21,7 град/мм [14]) обусловлено только дальним порядком его спиральных полимерных цепочек, мономерами которых являются ахиральные тетраэдры из молекул SiO2.

Рис 3. Температурные зависимости угла вращения (прибор ориентирован на восток;
точки – охлаждение; кружки – нагрев).
1 – физраствор + глюкоза (10%); 24.08.05.
2 – физраствор + глюкоза (40%); 28.09.05.
3 – физраствор + этанол (43%) + глюкоза
(25%); 05.09.05.
Очевидно, что в случае растворов сахаров энергия межмолекулярных взаимодействий (Е) будет определяться средним расстоянием между молекулами, то есть их концентрацией, а также их ориентацией относительно друг друга и геометрией надмолекулярных образований воды [2, 9]. При концентрации сахаров в растворах 20 – 40% (~2 моль/л) среднее расстояние между молекулами, равное ~10–7 см, сравнимо с их размерами. В этих условиях сахара должны ассоциировать, образуя упорядоченные надмолекулярные структуры под влиянием сил Ван-дер-Ваальса и при посредничестве динамичных водородных связей между молекулами воды [15]. Процесс ассоциирования будет сочетать в себе вращательную корреляцию самих сахаров с пространственной корреляцией включенных в ассоциат динамичных кластеров воды. Причем на обеих корреляциях будет сказываться хиральность электронной структуры молекулы сахара, в результате чего ассоциат и приобретает собственную хиральность, увеличивая тем самым суммарную оптическую активность раствора.
Таким образом, влияние температуры на динамику ассоциирования сахаров может обусловить температурную зависимость a их высококонцентрированных растворов. Суммарный эффект кооперации сахаров и молекул воды будет максимален при самой низкой температуре опыта, а значит, максимальным будет и значение a (обозначим его
a*). При повышении температуры степень корреляции и a понизятся в соответствие с распределением Больцмана:![]()
где Е – энергия взаимодействия или корреляции молекул. Из тангенса угла наклона линейной зависимости ln(
a/a*) = f(1/T) ?ычисляли энергию активации. Хорошая линейность данной зависимости была гарантирована малостью разницы (a* – a) по сравнению с a*. Наличие излома на некоторых температурных зависимостях при ~295К может быть связано с изломом в этой критической точке температурной зависимости динамической вязкости воды [8]. Этот результат подтверждает зависимость динамики кооперативных взаимодействий растворенных молекул от степени упорядоченности самой воды, которая возрастает по мере приближения температуры раствора к критическим точкам (4, ~22, ~36оС) [8]. О том же говорит увеличение величин Е при добавлении в растворы глюкозы и декстрана этанола (Рис. 3, 4; Таблица 1), молекулы которого могут оказывать стабилизирующий эффект на водородные связи в динамичных кластерах воды [9]. Известно [15], что ассоциирование стимулирует добавка NaCl и в согласии с этим величина Е для физраствора сахара больше, чем Е для его водного раствора (см. Таблицу 1).В работе [2] был зафиксирован локальный скачок величины
aО раствора ФР + ГК (0,8%) + сахар (20%) при неизменном значении aN около 5 часов утра 19.01.05, который явно не коррелировал с изменениями горизонтальных составляющих геомагнитного поля.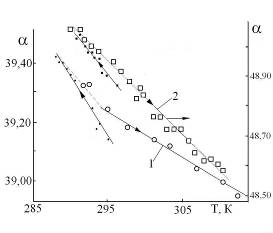
Рис 4. Температурные зависимости угла вращения (прибор ориентирован на восток; точки – охлаждение, кружки и квадраты – нагрев).
1 – физраствор + декстран (10%); 14.09.05.
2 – вода + сахар (~40%); 24.09.05.
Характер биогенного действия космофизических факторов в этот день в православной мифологии отмечен праздником “Богоявления” и таинством “водосвятия”. Как правило, в эти дни снижается температура (крещенские морозы) и, следовательно, возрастает порядок организации водных систем, в том числе и жидких сред организма человека. Очевидно, что метаболический эффект их упорядочивания оказывает заметное влияние на когнитивные функции мозга человека. В контексте данной гипотезы провели измерения a растворов ФР + глюкоза (10%) и ФР + декстран (10%) в период времени, на который приходится православный праздник Святого Духа (19.06.05) (Рис 5).
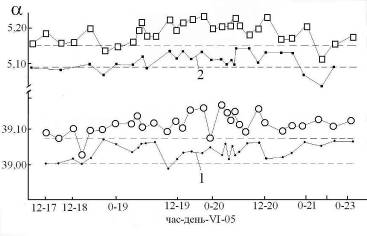
Рис 5. Зависимость угла вращения растворов от времени при Тком.
1 – ФР + декстран (10%) (кювета 200 мм);
2 – ФР + глюкоза (10%) (кювета 100 мм).
Ориентация прибора: точки – на восток, кружки и квадраты – на север. Измерения в комнате на восточной стороне дома. Пунктир – нулевая линия.
Ход зависимостей показывает увеличение a 19-го и 20-го 06.05, что можно связать с активизацией в эти дни хирального фактора космического масштаба. Обратный эффект – нивелировка хирального фактора – наблюдали во временной зависимости a раствора ФР + ХС (10%) + ГК (0,8%) + сахар (20%) в дни 12 – 15. 04.05 (Рис 6).

Рис 6. Зависимость угла вращения раствора ФР + ХС (10%) + ГК (0,8%) + сахар (20%) от времени при Тком. Ориентация прибора: точки – на восток, кружки – на север. Измерения от 8.04.05 до 2.04.05 в комнате на западной стороне дома, а 2 – 4.04.05 – в комнате на восточной стороне дома.
Наличие чувствительности
a и Da у этого раствора к возмущениям электромагнитных параметров биосферы может лежать в основе механизма метеозависимости людей, имеющих ту или иную патологию опорно-двигательной системы.В таблице 3 приведены величины энергий биогенных излучений и энергий активаций процессов, участвующих в энергоинформационном обмене живых систем с внешней средой. Величину ЕТР ~ QT для молекул воды оценили из зависимости коэффициента самодиффузии от температуры [16], для молекул сахаров эта энергия будет в три-четыре раза выше [17]. Соотношение Е < ЕВР, а также близость величин Е для глюкозы, сахара и декстрана, для которых мутаротация маловероятна [1], позволяют исключить влияние мутаротации на температурные зависимости a глюкозы.
Таблица 3.
ВЕЛИЧИНЫ ЭНЕРГИЙ АКТИВАЦИИ И БИОГЕННЫХ МКВ-КВАНТОВ
Природа и характеристика кванта энергии |
Обозначение |
Величина (кДж/моль) |
Энергия активации реакции ассоциирования Сахаров |
Е |
0,1 – 0,6 |
Тепловая энергия (kТ) |
QT |
2,5 |
Тепловая критическая (k |T – Tкр|) |
Q DT |
~0,1 |
Активация трансляционной подвижности молекул воды [18] |
ЕТР |
~2 |
Крутильные колебания больших молекул в конденсированной фазе (w ~ 1011 с–1 [17]) |
ЕW |
~0,04 |
Барьеры внутримолекулярного вращения |
ЕВР |
~ 4,0 |
Энергия резонансного возбуждения кластеров воды ( lрез ~ 6 мм [20]) |
ЕРЕЗ |
0,02 |
Биогенное МКВ-излучение ( l = 100 – 1 мм [21]) |
ЕМКВ |
0,001 – 0,1 |
Реликтовое излучение ( l ~1 мм) |
ЕR |
0,1 |
Энергия нейтринных флуктуаций вакуума [6, 23] |
ЕО |
~10–16 |
Известно [8, 20], что в окрестностях критической точки фазового перехода роль упорядочивающего фактора играет разность температур |Т – Ткр|. Эти данные и то, что величина Е, как правило, существенно больше значений Q
DT, ЕW, ЕРЕЗ, ЕМКВ и ЕR, но на порядок меньше QT и ЕТР, позволяют принять способность сахаров образовывать хиральные ассоциаты в физиологических жидкостях за основу механизма адаптации живых систем к изменениям внешних физических факторов. Действительно, вода и водные растворы метаболитов резонансно поглощают слабое МКВ-излучение (ЕРЕЗ), которое может активизировать перестройки надмолекулярных структур [18, 19]. Например, при МКВ-облучении (l = 10 см) водного раствора гликогена раствор утрачивал оптическую активность [19]. Учитывая величину ЕR и гипотезу о биогенности нейтрино [21], предположим, что реликтовое излучение и солнечное нейтрино, как постоянно действующие факторы биогенеза, внесли свою лепту в развитие механизма адаптации или энергоинформационной связи живых систем с внешней средой. Реликтовое излучение, например, может быть ответственно за механизм и частоты спонтанной пульсации пейсмекеров вегетативной нервной системы.Упорядочивающий эффект фазового перехода в хиральной живой системе, очевидно, сопряжен с резонансной синхронизацией крутильных колебаний молекул и последующей конденсацией квантов энергии порядка ЕW, ЕРЕЗ, ЕМКВ и ЕR в квант энергии порядка энергии макроэргической связи. В принципе, по такому же механизму кванты энергии порядка ЕО могут конденсировать в кванты Q
DT и ЕR [21] в момент пространственной корреляции однотипных электронных орбиталей в молекуле полисахарида, имеющей в своем основании гиалуроновую кислоту (Рис 7). Очевидно, что такие переходы идут в информационных системах живого организма, имеющих однородно множественную молекулярно-клеточную структуру, предрасположенную к гелеобразованию.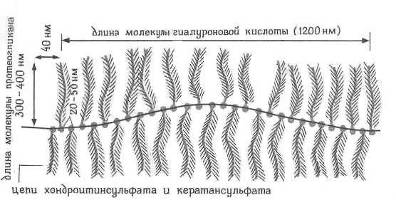
Рис7. Схема макромолекулы, играющей активную роль в энергетике соединительной ткани [11].
К таким системам относятся, например, стекловидное тело глаза, цитоплазма аксона, синовия. Сами же фазовые переходы в информационных системах наиболее вероятны в состоянии сна, когда снижается температура организма. По-видимому, в основе парадоксальной фазы сна, характеризуемой быстрым движением глаз (фаза-БДГ), тоже лежат фазовые переходы, обеспечивающие корректировку гомеостаза внешними и внутренними хиральными факторами.
Результаты настоящей работы позволяют заключить, что предрасположенность физиологических жидкостей содержащих метаболические сахара к фазовым переходам в критических температурных точках воды, может лежать в основе механизма адаптации живых систем к изменениям внешних физических условий, как в геологическом, так и в реальном масштабе времени.
ЛИТЕРАТУРА
1. Кульневич В.Г. // Соросовский образовательный журнал. – 1996. – № 8.
2. Холманский А.С. // Математическая морфология. Электронный журнал. – Т. 5. – Вып. 3. - 2005. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/N-11-html/cholmansky/cholmansky.htm3. Кизель В.А. Физические причины диссимметрии живых систем. – М.: “Наука”, 1985.
4. Жвирблис В.Е. // Космофизические истоки дисимметрии живых систем. – М. 1987.
5. Пармон В.Н. // Вестник РАН. – 2002. – Т. 72. – № 11.
6. Холманский А.С., Стребков Д.С. // Доклады РАСХН. – 2004. – № 1.
7. Холманский А.С. http://www.sciteclibrary.ru/rus/catalog/pages/7876.html
8. Холманский А.С. // Доклады РАСХН. – 2006. – № 2.
9. Смирнов А.Н. и др. // Электронный журнал “Исследовано в России”
http://zhurnal.ape.relarn.ru/articles/2004/038.pdf 413
10. Досон Р. и др. Справочник биохимика. – М.: “Мир”. 1991
11. Мусил Я., Новакова О., Кунц К. Современная биохимия в схемах. – М.: “Мир”. 1984
12. Холманский А.С. “Способ определения функционального состояния человека”. Патент РФ 2193859 от 10.07.2001. Бюл. 10.12. 2002. – № 34
13. Жвирблис В.Е. // Изв. АН СССР. Сер. биол. – 1982. – № 3.
14. Физические величины. Справочник. – М.: Энергоатомиздат. 1991.
15. Кизель В.А. Практическая молекулярная спектроскопия. – М.: МФТИ. 1998
16. Севрюгин В.А., ЖуравлеваН.Е., Смирнова Л.Г. Химия и компьютерное
моделирование. Бутлеровские сообщения
.
2002, № 617. Кобозев Н.И., Избранные труды. Т. 2. М.: МГУ. 1978
18. Синицын Н.И. и др. // Биомедицинская радиоэлектроника. –1998. – №1.
19. Кудряшов Ю.Б. Биофизические основы действия микроволн. – М.: МГУ. 1980.
20. Дайсон Ф. и др. Устойчивость и фазовые переходы. – М.: “Мир”. 1973
21. Холманский А.С. // http://www.sciteclibrary.ru/rus/catalog/pages/6303.html
Dependence of Optical Activity of Physiological Solutions of Sugar on Temperature
Holmansky A.S.
GNU UNII of an electrification of an agriculture, Moscow
Temperature dependence of optical activity of physiological solutions of metabolic sugars is investigated and energy of activation of reaction of assosiation sugars in supramolecular chiral structures is determined. Proceeding from value of energy of activation and taking into account influence on optical activity of time, a place and orientation of the device have stated the assumption of a key role of sugars in the mechanism of adaptation of alive systems to changes chiral factors of an environment.
ГНУ ВНИИ электрификации сельского хозяйства, Москва
Холманский Александр Сергеевич (asholman@mtu-net.ru)
Поступила в редакцию 28.12.2005.