УДК 515.015
ДИХОТОМИЯ ПРАВОГО И ЛЕВОГО
Ó
2007 г. Холманский А. С.
Анализ структурных
особенностей водных кластеров и метаболитов (сахаров, АТФ, ДНК, РНК, гормонов)
показал универсальность тетраэдрической метрики электронных орбиталей. Для тетраэдров с асимметричным центральным
атомом, имеющим нулевой ядерный спин
(С, О, S) предположили
взаимодействие валентных электронов с внешним хиральным фактором
электромагнитной природы, а для ядер с ненулевым спином (Р, J) к данному взаимодействию добавили еще спин-спиновое взаимодействие
между хиральным квантом и ядром. Второй тип взаимодействия позволяет привлечь
гипотезу хирального фактора нейтринной природы для объяснения причин дисимметрии
биосферы, начиная с пребиотического этапа эволюции. Совокупное действие двух
хиральных факторов на живые системы отвечает за генезис
функционально-морфологической асимметрии правого и левого полушарий, лежащей в
основе физики мышления.
Ключевые слова: дихотомия, дисимметрия,
хиральность
Предельно сложные формы
дихотомии реализуются в иерархии живых систем. К ним можно отнести симбиоз
мужских и женских организмов, синергизм правого и левого полушарий мозга
человека и даже обрядово-догматические различия у монотеистических религий [1,
2]. Своеобразна и требует изучения дихотомичность метаболизма, который сочетает
физико-химические свойства воды, правых (D) сахаров и левых (L)
аминокислот. Философская база парадигмы дихотомии зиждется на «хирально
стерильных» законах диалектики (закон подобия, единства и борьбы
противоположностей). Они не содержат правил комбинирования дискретных форм
материи, обусловивших асимметрию между право- и левовинтовым движением на
уровне элементарных частиц. Поэтому открытыми остаются вопросы: почему процесс
самоорганизации мира уже в самом начале предпочел частицы, а не их зеркальные
антиподы – античастицы? Существует ли в природе
универсальный физический фактор, под действием которого мир, а затем и биосфера
приобрели диссиметрию и хиральность [3]?
Очевидно, что данный фактор сам должен быть
хиральным, кроме того, живые
водосодержащие системы в отличие от косных хиральных структур должны
обладать специфическим механизмом реагирования на действие такого фактора. Л.
Пастер, открыв явление оптической активности в 1860 году, предположил, что
асимметрия биосферы есть результат действия электрических и магнитных полей
космического происхождения. В развитие этой гипотезы В. Вернадский связал
хиральность живых систем с асимметрией самого пространства. Обе гипотезы
разумны и дополняют друг друга. Известно [4], что природные электромагнитные
поля, как правило, асимметричны, частично поляризован солнечный свет. Есть
работы [4-6], в которых обсуждаются модели механизма чувствительности живых
систем к хиральному фактору, основанные на хиральности электромагнитного поля,
структурной асимметрии метаболитов и кооперативных явлениях в водных
атомно-молекулярных системах.
Изучать
природу хирального фактора можно двумя
способами.
1.
Исследовать
хиральные свойства универсальных физических факторов эволюции. На их роль могут
претендовать хиральные поля и кванты электромагнитной и нейтринной природы.
2.
Изучать
зависимость физико-химических свойств водных растворов метаболитов от их структуры и внешних условий.
В настоящей работе, в качестве модели хирального кванта по
примеру Дж. Максвелла [7] использовали вихрь идеальной сплошной среды,
сочетающий в себе вращательное и поступательное движения (Рис. 1). В
зависимости от направления векторов импульса и момента импульса вихрь может
быть право и левоспиральным. Такой вихрь можно взять за основу кванта энергии,
как электромагнитной (ЭМ), так и нейтринной природы. Кинематика и динамика
ЭМ-вихря определяются правилом Ленца и правым буравчиком Максвелла [1, 7].
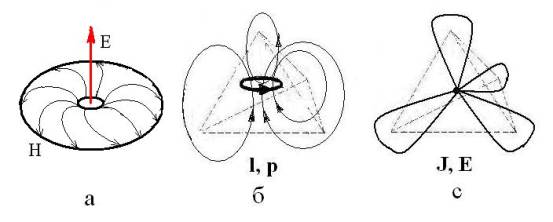
Рис. 1. Модели хирального
ЭМ-вихря [10] (а), хирального кванта с моментом импульса – l и
импульсом – р (б) и схема sp3-гибридизации валентных
электронов атома. J – ядерный спин, Е, Н – напряженности электрического
и магнитного поля.
Универсальным элементом геометрии пространств литосферы,
гидросферы Земли и водной среды жидких систем является тетраэдр, отвечающий sp3-гибридизации межатомных или межмолекулярных связей (Рис.
1, 2). Это, прежде всего, относится к содержащим кварц верхним слоям земной
коры, пескам и глинам. Высокая доля кремнезема (SiO2) в этих средах располагает
к формированию сеток из цепочек центрированных тетраэдров (Рис. 2). В
низкотемпературной модификации кварца цепочки тетраэдров располагаются по
спирали [8], поэтому прозрачный кварц оптически активен.
Элементом трехмерных сеток водородных связей в конденсированных
водных системах также является тетраэдр (Рис. 2). В работе [9] предположили,
что в воде возникают короткоживущие надмолекулярные образования (кластеры) изоморфные
цепочкам тетраэдров в кварце, благодаря которым вода может приобретать
динамическую оптическую активность. Стационарная концентрация хиральных
кластеров воды должна определяться их временем жизни и, очевидно, возрастает
при растворении в воде хиральных молекул, предрасположенных к образованию
водородных связей и ассоциированию (самоорганизации). Такими молекулами
являются аминокислоты, белки и сахара. Синергизм динамической оптической
активности воды и внешнего хирального фактора, может избирательно влиять на
кинетику ферментативных реакций синтеза хиральных метаболитов, например
гормонов роста и цветения [10]. По-видимому, в этом механизме свою роль играют
и хиральные среды литосферы, которые могут селектировать и концентрировать
потоки хиральных квантов, как исходящих из недр Земли, так и космической природы.
В основе
энергоинформационного действия хирального фактора должен лежать механизм
взаимодействия хиральных квантов с многокомпонентными водными средами живых
систем. Ядерно-электронная структура, участвующая в данном взаимодействии,
вероятнее всего будет иметь геометрию тетраэдра. Простая трансформация вихревой
модели хирального кванта может обеспечить механизм его резонансного поглощения
ядерно-электронной структурой атома или молекулы метаболита (Рис. 2). Симметрия
sp3-гибридизации является производной внутренней
структуры ядра, которая задает число и порядок формирования электронных
оболочек атома, в том числе и геометрию орбиталей валентных электронов.
Тетраэдрическую стереохимию метаболитов и их фрагментов определяют помимо ядер
С12 (0), С13 (1/2) такие ядра: Р (1/2), S (0), Cl
(3/2), J (5/2) (в скобках приведены значения ядерных спинов). В случае различия
орбиталей всех четырех связей данных атомов они становятся центрами асимметрии,
наделяя соответствующие структуры
свойством хиральности.
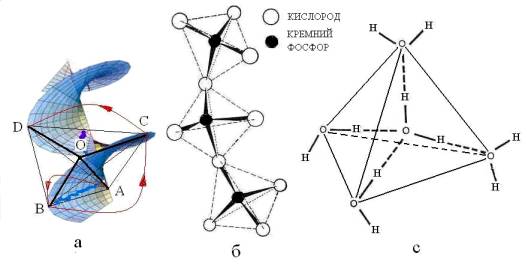
Рис. 2. Спиральная метрика
пространства (синий цвет), в которую вписан тетраэдр ABCD. Красные стрелки – вращения
радиус-вектора ОА против часовой стрелки моделирует один оборот правого винта
спирали. Цепочка тетраэдров SiO4 в кварце или РО4
в воде (б). Тетраэдрический элемент надмолекулярной структуры воды (с).
В случае если хиральный
квант есть ЭМ-вихрь, его поглощение осуществляется в рамках взаимодействия с
валентными электронами. При этом спин ядра и его магнитный момент не играют
какой-либо роли. Очевидно, что метаболиты с асимметричными атомами углерода (С12)
и серы будут чувствительны к хиральному фактору электромагнитной природы
(ЭМ-фактор), поскольку их ядерный спин равен нулю. Область влияния ЭМ-фактора
охватывает все процессы производства и расходования в организме
электромагнитной энергии, источником которой являются ферментативные реакции
окисления углеводов, белков и жиров. Электромагнитная энергия обеспечивает
работу нервов, мышц и половых органов.
При окислении правой глюкозы
образуется в целом оптически не активная молекула АТФ. Однако при анаэробном
гликолизе глюкозы помимо двух ахиральных молекул АТФ образуются две молекулы
хиральной молочной кислоты, имеющей тетраэдрическую структуру. Ее роль в
метаболизме отнюдь не исчерпывается энергетической функцией. К примеру, в
работе [1] высказана гипотеза, что синтез молочной кислоты в стекловидном теле
глаза асимметричен, то есть в правом глазу преобладает синтез, допустим, L-изомера,
а в левом – D-изомера. Косвенно эта гипотеза подтверждается тем,
что слова русского языка лучше
распознаются в правом поле зрения или левым полушарием мозга, а слова
еврейского языка (Рис. 3) – в левом поле зрения и, соответственно, в правом
полушарии мозга [11]. Если сравнить графику букв этих двух языков, то можно
обнаружить, что в иврите преобладает элемент зеркально симметричный русской
букве «Г» (Рис. 3). На основании этих данных можно заключить, что
ферментативный синтез молочной кислоты в правом и левом глазе зеркально
симметричны и эта асимметрия может передаваться также на мозг, поскольку
жидкость стекловидного тела близка по составу с ликвором и молочная кислота
может диффундировать по зрительному нерву в мозг. Таким образом, молочная кислота,
являясь носителем ЭМ-фактора, может привносить соответствующую асимметрию в
электромагнитную энергетику мозга и всего организма.
Рис. 3. Фрагмент текста на иврите
Регуляторами
электромагнитных процессов ответственных за половую функцию организма являются
стероидные гормоны (Рис. 4), химическая структура которых вполне может
обеспечить аккумуляцию квантов электромагнитной природы с тетраэдрической
метрикой. Аналогичную роль может исполнять в мозгу химический предшественник
половых стероидов – холестерол. А в растениях эту роль исполняют гормоны роста
и цветения (например, гиббереллин) [9], имеющие родственную со стероидами
структуру.
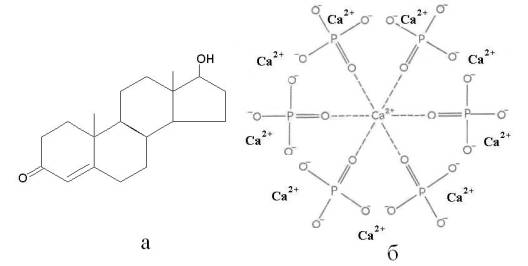
Рис. 4. Структуры полового
гормона тестостерона (а) и молекулярной основы костного материала – кристалла гидроксиапатита (б).
Предположим, что поглощение
кванта нейтринной природы сочетает в себе резонансное взаимодействие магнитных
моментов (спинов) кванта и ядра, а также импульса кванта с электромагнитным
полем валентных электрона. Последнее взаимодействие, как и в случае хирального
ЭМ-фактора, будет приводить к химической активации хромофора или к повышению
химического потенциала метаболита [1]. Взаимодействие же спинов кванта и ядра,
очевидно, будет менять ориентацию последнего. Данное взаимодействие аналогично
явлению ЯМР и взаимодействию магнитных моментов ядер атома водорода и
диамагнитного тока электронов в молекулах биосистемы с внешними полями
напряженностью 10–9 – 10–12 Тл [12]. Таким образом, можно
полагать, что хиральный фактор нейтринной природы (J-фактор) играет важную роль
в кооперативных явлениях и в механизмах энергоинформационного обмена. Конечно,
в силу малости энергии хиральных ЭМ- и J-факторов их
электромагнитное и спин-спиновое взаимодействия с метаболитами будут иметь характер
возмущений.
Детали механизма действия J-фактора
на живые системы можно узнать из анализа свойств двух биогенных элементов –
четырехвалентного фосфора и йода, валентность которого меняется в пределах от
одной до восьми [8]. Фосфор в живых системах и в естественных водоемах
присутствует в виде аниона фосфата РО4–, имеющего
геометрию тетраэдра и образующего цепочки подобные, представленной на Рис 1.
Фосфат присутствует в метаболитах, играющих ключевую роль в энергетике и
передачи информации в живом организме. К ним относятся: молекула-аккумулятор
электромагнитной энергии (АТФ), ДНК, РНК (Рис. 5); а также кристалл
гидроксиапатита – основа костной материи
(Рис. 4).
Йод входит в состав важного гормона щитовидной железы –
тирозина, который повышает активность гормонов роста и многих ферментов, прежде
всего, ферментов расщепляющих углеводы, он также отвечает за умственное
развитие в раннем детском возрасте. Недостаток йода у детей ведет к развитию
идиотизма, а у взрослых – к замедлению всех метаболических процессов. В
организме выработался механизм поддержания концентрации йода в крови на одном
уровне. Наблюдаются сезонные вариации концентрации йода в крови: снижение с
сентября по январь и достижение максимального значения в мае – июне. Эти
вариации могут быть связаны с ритмикой биогенного излучения Солнца,
ответственного за размножение живых организмов.
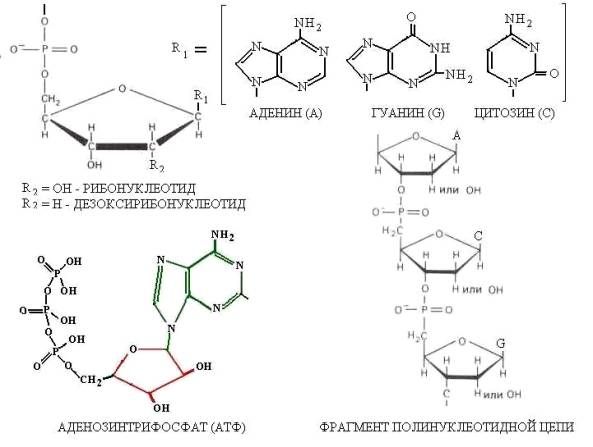
Рис. 5. Структуры биологических молекул содержащих
тетраэдрический фосфор
Первичным источником биогенного фосфора и йода является гидросфера
Земли. В трофическую цепь эти элементы вовлекаются в процессе фотосинтеза
фитопланктона. Учитывая, что вода в значительной мере экранирует действие
хирального ЭМ-фактора, в ней возрастает роль хирального J-фактора, аккумулятором которого могут
выступать тетраэдрический фосфор и йодсодержащие органические соединения.
Данное свойство ядерно-электронной структуры фосфора и йода могло
предопределить их функции в метаболизме и заложить уже на пребиотическом этапе
эволюции основные принципы структурной и функциональной асимметрии живых
систем.
Структуры АТФ, ДНК и РНК имеют в своей основе один и тот же
нуклеотид, состоящий из хирального сахара (D-рибоза), азотистого
основания и фосфатной группы, связанной с рибозой через СН2-мостик
(Рис. 5). Такой нуклеотид, в принципе, может аккумулировать на своей
ядерно-электронной структуре хиральные кванты, как электромагнитной
(ЕМ-фактор), так и нейтринной (J-фактор) природы.
Структурное подобие нуклеотидов АТФ, ДНК и РНК способствует эффективному обмену
между ними квантами соответствующей энергии по резонансному механизму. Такой
обмен может ускорять и хирально дифференцировать кинетику ферментативных
реакций, обеспечивающих процессы отбора и передачи генной информации (трансляция РНК, репликация и транскрипция
ДНК). Высокое содержание фосфата в
костной ткани и симметрия кристаллов гидроксиапатита будут способствовать
аккумуляции хиральных квантов нейтринной энергии, которая может принимать
непосредственное участие в процессе кроветворения в костном мозге.
Таким образом, проведенный анализ структурных особенностей
водных растворов метаболитов показал универсальность тетраэдрической метрики
электронных орбиталей. Для структурных тетраэдров с асимметричным атомом,
имеющим нулевой ядерный спин
предположили взаимодействие валентных электронов с внешним хиральным
фактором электромагнитной природы, а для ядер с ненулевым спином к данному
взаимодействию добавили еще спин-спиновое взаимодействие между моментом
импульса хирального кванта и спином ядра. Второй тип взаимодействия позволяет
привлечь гипотезу хирального фактора нейтринной природы для объяснения причин
дисимметрии биосферы. Совокупное действие двух хиральных факторов на живые
системы охватывает весь диапазон функционально-морфологической асимметрии
организма человека.
ЛИТЕРАТУРА
1. Холманский А. С.
Моделирование физики мозга // Математическая морфология. Электронный
математический и медико-биологический журнал. - Т. 5. - В. 4. - 2006. – URL:
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky-3/holmansky-3.htm
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/TITL-12.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/cont.htm
2. Ресурс функциональной
асимметрии мозга //Математическая морфология. Электронный математический и
медико-биологический журнал. -Т. 5. - Вып. 4. - 2006. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky-4/holmansky-4.htm
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/TITL-12.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/cont.htm
3. Кизель В. А. Физические причины диссимметрии живых систем. – М.: «Наука». 1985.
4. Жвирблис В.
Е. Космофизические истоки дисимметрии живых систем // Принципы симметрии и
системности в химии, изд-во Московского ун-та, 1987 г., 123 С.
4. Холманский А. С.
Зависимость от температуры оптической активности физиологических растворов
сахаров //Математическая морфология. Электронный математический и
медико-биологический журнал. – Т. 5. – Вып. 4. - 2006. – URL:
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky/holmansky.htm
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/TITL-12.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/cont.htm
5. Юхновский Ю.Р. Симметрия и фазовые переходы второго рода. В сб. Методологические проблемы кристаллографии. М.: Наука. – 1985. 296 с.
7. Максвелл
Дж. О физических силовых линиях // Избранные очинения по теории
электромагнитного поля. – М.:. 1954. - С.105 – 248.
8. Пиментол П., Спратли Р. Как
квантовая механика объясняет химическую связь.
- М.: Мир, 1973э
9.
Холманский А. С. Термодинамические
особенности воды и биоэнергетика //Доклады РАСХН, 2006. 2. //
http://www.sciteclibrary.ru/rus/catalog/pages/7897.html
10. Желудев И. С. Симметрия и ее приложения. - М.:
Атомиздат. 1976.
11.
Физиология человека. Ред. Р. Шмидт, Г. Тевс, Т. 1-4, - М.: Мир, 1996.
12. Пушкарев Ю.
П. Удивительная физиология. – СП., 2005.
- С. 104
13. Леднев
В. В. Биоэффекты слабых комбинированных, постоянных и переменных
магнитных полей. // Биофизика. 1996. Т. 41, № 1. С. 224-232.
Dichotomiy of
the right and left
Kholmanskii A. S.
The analysis of
structural features water clasters and metabolitics (sugars, ATP, DNA, RNК, hormones) has shown universality
tetrahedral metricss electronic orbitals. For tetrahedrons with the asymmetric
central atom having zero nuclear backs (C, O, S) have assumed interaction
valent electrons with external chiral the factor of the electromagnetic nature,
and for nuclear with nonzero spin (Р, J) to the given interaction have added still
backs-spin interaction between chiral quantum and a nuclear. The second type of
interaction allows to involve a hypothesis chiral the factor neutrino the
nature for an explanation of the reasons disymmetric biospheres, since
prebiotical a stage of evolution. Cumulative action of two chiral factors on
alive systems is responsible for genesis of is functional-morphological
asymmetry of the right and left hemispheres underlying physics thinking.
GNU UNII of
an electrification of an agriculture, Moscow
ГНУ ВНИИ электрификации
сельского хозяйства, Москва
Поступила в редакцию 2.05.2007.