- Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 7. -
Вып. 1. - 2008. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-17-html/TITL-17.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-17-html/cont.htm
УДК:591.471.321+591.48+591.192
Собственная гипотеза происхождения позвоночных и их симметрии,
сегментации, хорды и трубчатой
нервной системы*
Ó 2008 г. Ермоленко А. Е. Перепада Е. А.
Предложена новая гипотеза происхождения позвоночных.
Развитие позвоночных начинается с образования многоклеточного организма путем
упорядоченного многократного деления репродуктивной клетки и не расхождения
вновь образовавшихся клеток, сохранивших между собой связь с помощью
внеклеточного матрикса. Далее происходит последовательное образование организмов за счет агрегации себе подобных структур. Таким
способом образовались: бластея; гастрея
Э. Геккеля; усовершенствованная гастрея (супергастрея), организм
наподобие ползающего гребневика; сегментелла Yermola, состоявшая из небольшого
количества (2-9) супергастрей, организм наподобие олигомерных червей;
суперсегментелла, образованная путем соединения задних концов двух сегментелл.
Суперсегментеллы дали развитие оболочникам, полухордовым, хордовым типа
ланцетника и предшественникам позвоночных. Происхождение органов: от внеклеточного матрикса
первых многоклеточных произошла
мезоглея, а затем и хорда; энтодермальные впячивания дали развитие первичной
кишке; эктодермальные впячивания после сосредоточения там ранее диффузно расположенных
нервных клеток преобразовались сначала в желоб, а затем в трубчатую нервную
систему; уплотненная нервная ткань аборальных
полюсов дала развитие узловой нервной системе. Железистые клетки
супергастреи явились производным материалом для всех желез в соответствии с
первичной сегментацией.Сегментация организмов обусловлена агрегацией
супергастрей в единый организм при двухплоскостной симметрии. Происхождение
симметрии: симметрия обусловлена структурообразующим свойством внеклеточного
матрикса. Симметрия первичных организмов была радиальной; затем
радиально-билатеральной, при этом первая
плоскость симметрии разделяла организм на дорсальную и вентральную
стороны. С расположением супергастрей в ряд последовательно создалась
радиально-двухплоскостная симметрия, при этом новая сагиттальная плоскость
разделяла организм на правый и левый антимеры. При объединении двух организмов
(сегментелл) задними концами образовалась радиально-трехплоскостная симметрия.
Третья плоскость симметрии разделяла организм на передний и задний антимеры.
Ключевые слова: хордовые, позвоночные, симметрия, нервная система,
гастрея, внеклеточный матрикс, первичная кишка.
Краткий обзор сведений о происхождении позвоночных
Происхождение позвоночных –
нерешенный вопрос биологии. Путем сопоставления строения и развития животных и
собственных предположений сравнительные анатомы разработали множество концепций
происхождения Bilateria, в том числе позвоночных. Сведения об этих концепциях с
их анализом можно найти в монографиях [10, 19, 66], диссертациях [5] и обзорных
статьях [14, 28, 41, 44, 45]. Одна из первых концепций для решения этого
вопроса была предложена Сент-Иллером Ж. [22] еще в первой половине XIX
века. Он считал, что позвоночные могут рассматриваться как перевернутые
членистоногие. Сходную идею о существовании предков хордовых, похожих на
кольчатых червей, развивал Дорн А. [6]. И в XXI веке эта идея жива и имеет
своих сторонников [14, 32, 42, 50, 57]. Сущность идеи заключается в том, что
расположение сердца и направление кровотока у хордовых и членистоногих, если
последних перевернуть, совпадает. Также в этом случае совпадает расположение
целомов. На этом основании сторонники этой идеи считают, что предки хордовых в
своем эволюционном развитии испытали инверсию сторон тела, то есть
перевернулись и стали передвигаться на морфологически спинной стороне, которая
стала функционировать как физиологически брюшная, а морфологически брюшная
сторона предков хордовых стала функционировать как физиологически спинная.
Направление кровотока у кольчатых червей, членистоногих и хордовых (если
последних перевернуть) совпадает. Однако, возможно, что разное расположение
сердца (снизу у хордовых и сверху у беспозвоночных) связано с независимым
возникновением у них кровеносной системы.
Сторонники гипотезы
перевернутости хордовых указывают, что некоторые современные виды ведут
перевернутый образ жизни (вниз спиной). Однако остается факт, что при этом
спинная и брюшная стороны остаются таковыми вне зависимости от перевернутости.
Известно, что у «неправильных» гребневиков (Ctenophora) ротовое и/или анальное
отверстия смещены, однако не вызывает сомнения энто- и эктодермальное
происхождение этих отверстий. В
доказательство правильности этой гипотезы ее сторонники приводят данные
молекулярной биологии [2, 29]. У зародышей некоторых
позвоночных на стадии гаструлы
на брюшной стороне синтезируется белок BMP-4 (bone morphogenetic
protein-4), а на спинной - белок CHD (chordin). В норме белок CHD синтезируется
на спинной стороне, но если в опыте повысить его концентрацию на брюшной с
помощью инъекции, то на этом месте в дальнейшем разовьются структуры спинной
стороны. То же происходит с искусственным повышением на спинной стороне
зародыша концентрации белка BMP-4, который вызывает развитие там структур брюшной стороны. Аналогичные
результаты были получены и на дрозофиле. Сходные данные также были получены Slack J. с
соавторами [61] при исследовании генов Hox-группы в развитии различных
беспозвоночных и позвоночных животных. В этих публикациях было показано, что у
беспозвоночных эти гены маркируют брюшную сторону, а у позвоночных их
экспрессия наблюдается на спинной стороне. Однако,
нахождение белков там, где они есть, не подтверждает, но и не опровергает
гипотезу перевернутости, а только указывает, что эти белки формируют
определенные структуры или как-то влияют на их формирование и не более.
Вероятно, эти белки являются производными индуктора бластопора (организатора
развития организма) при ранней гаструляции.
Сторонников этой идеи не
останавливает наличие значительных расхождений в развитии и устройстве этих
групп животных, отсутствие гомологичных органов. Первично- и вторичноротые
отличаются характером дробления оплодотворенного яйца: у первичноротых оно
спиральное, а у вторичноротых - радиальное. Другое отличие состоит в способе
закладки целома: у первичноротых стенки вторичной полости тела происходят от
двух телобластов, а у вторичноротых - за счет выпячивания карманов
эмбрионального кишечника. Важнейшим различием между хордовыми и членистоногими
является устройство центральной нервной системы. У членистоногих и аннелид
отсутствует трубчатая нервная система, и наличие ее у хордовых нельзя объяснить
переворачиванием.
Таким образом, гипотезы,
объясняющие план строения хордовых и их происхождение от перевернутых аннелид
или членистоногих, не могут быть признаны корректными в связи с тем, что
имеются фундаментальные эмбриологические отличия между кольчатыми червями и
членистоногими, с одной стороны, и хордовыми - с другой.
Рассматривались гипотезы
происхождения хордовых от кишечнодышащих. У этих животных (как и у хордовых)
имеются жаберные щели, а в передней части тела - опорный вырост кишечника
(стомохорд), который сравним с хордой. Однако кишечнодышащие резко отличались
от хордовых обратным направлением тока крови, расположением сердца, строением
нервной системы и другими важными особенностями организации. Делались попытки
выводить хордовых от иглокожих, немертин, моллюсков и других групп
беспозвоночных, однако ни одна из них не имела успеха [1, 8, 9, 48, 49, 51, 53,
54].
Некоторые авторы гипотез
выводят происхождение позвоночных из древних форм современных полухордовых. Но
авторы, которые приводят факт сходства полухордовых и хордовых, не могут
установить степень их родства и указать
пути превращения первых во вторых. В дополнение к этой идее существуют
неотенические гипотезы [1, 30, 60]. В
соответствии с некоторыми из них [30] хордовые произошли от личинок древних
асцидий, имевших хорду и нервную трубку. Более вероятно, что эти особенности -
наследие свободноживущих предков асцидий, о происхождении и организации,
которых неотенические гипотезы ничего не говорят.
Обстоятельный анализ существующих гипотез о
происхождении хордовых сделан Савельевым С. В. [19]. Этот анализ позволяет ему
сделать обоснованный вывод о несостоятельности
существующих теорий о происхождении
хордовых, с чем мы полностью согласны.
Что касается предложенной им собственной гипотезы происхождения нервной системы
хордовых, фактически самих хордовых, то она заслуживает обсуждения, и мы с ней не можем полностью согласиться.
На основании того, что палеонтологическая летопись содержит очень скудные
данные о происхождении хордовых и нет сведений о причинах, которые привели к
появлению первых хордовых, автор приходит к выводу, что все современные теории построены
на весьма «сомнительных предположениях», и в связи с этим «имеют право на
существование самые невероятные гипотезы эволюционного происхождения
позвоночных». Савельев С. В. выдвигает идею о происхождении хордовых от
гипотетического плоского животного с двумя парами нервных тяжей, которое из-за
изменения структуры морского грунта повернулось на 90°. Так образовались новые
брюшная и спинная поверхности. Сам автор идеи очень сомневается в вероятности
такого события, поэтому высказывает еще одну идею о происхождении трубчатой
нервной системы, а следовательно и хордовых, от древних турбеллярий, имеющих
родственные отношения с современными турбелляриями (Aloeocoela).
Надо отметить, что автор
идеи происхождения хордовых от древних турбеллярий пытается решить вопрос о
происхождении трубчатой нервной системы. По его мнению, образование нервных
тяжей обеспечивается функционированием организма. При зарывании его в ил
происходило трение и смещение нервных клеток, что приводило к слиянию ганглиев. Савельев С.В. находит оригинальное решение в
проблеме происхождения нервной трубки - центральная нервная система хордовых
произошла из слившихся дорсальных нервных стволов древних турбеллярий. В
результате объединения спинных ганглиев сформировалось протяженное скопление
нервных клеток, неразделенное участками нервных волокон. В связи с тем, что
типичный ганглий беспозвоночного состоит из клеток, находящихся на его
периферии, а центральная часть состоит из переплетенных отростков нервных
клеток, и в связи с потребностью в циркуляции жидкости вокруг нейронов
образовалась полость внутри нервного тяжа. Так образовалась трубчатая нервная
система. Нам представляется, что нервная система первых хордовых действительно
имеет две пары нервных тяжей, которые сегментировались и присутствуют у всех
современных позвоночных в виде узлов периферической нервной системы. Таким
образом, эти тяжи не могли быть использованы для образования трубчатой нервной
системы.
По мнению Савельева С.В.
[19] хорда появилась в связи с потребностью организма в специальной несущей
конструкции, так как образовавшаяся нервная трубка располагалась между двумя
тяжами мускулатуры, и организму в отсутствие такого каркаса было бы невозможно
поддерживать стабильную форму и нормальную циркуляцию спинномозговой жидкости.
На наш взгляд, такое объяснение нерационально. Во-первых, потому, что органы не
появляются в связи с какой-либо потребностью организма. Имеющиеся органы могут
использоваться организмом для выполнения каких-либо дополнительных
функциональных нагрузок. Во-вторых, из эмбриологии известно, что хорда
появляется раньше нервной трубки [11], и удаление хорды в эксперименте приводит
к развитию эпидермиса вместо нервной ткани [63, 64]. Появление жабр автор также
объясняет потребностью организма, находящегося в новых условиях зарытым в грунт
для поддержания повышенного метаболизма. Своеобразно автор новой гипотезы
происхождения хордовых решает вопрос о происхождении билатеральной симметрии.
Он считает, что в связи с переходом свободноплавающих животных к придонному образу
жизни появились 4 нервных ствола, а в результате этого возникла первая ось
симметрии, т. е. он пытается объяснить появление симметрии функцией организма и
образом жизни. Вывод такой не вполне корректен, так как симметрия
биологического организма, также как и живого минерального организма
(кристалла), обеспечивается их внутренним составом [25]. Также не обоснованы
попытки автора объяснить сегментацию, основанную на метамерии мускулатуры,
которая появляется в связи с
потребностью в движении.
Нам импонирует идея,
изложенная Дьюел Р [37], в основе которой лежит предположение, что
двухсторонесимметричные животные произошли в результате интеграции колоний
кишечнополостных особей. Такие взгляды соответствуют ранее предложенной нами
гипотезе [38, 39]. Однако мы не разделяем мнение автора, что метамерно
расположенные полипы колонии превратились в отдельные органы
билатерально-симметричного индивидуума. На наш взгляд произошла интеграция не
простых двухслойных гастрей, а высокоорганизованных индивидуумов, имевших экто-, энто- и мезодерму, а также мезоглею,
нервную, железистую ткани и половые клетки.
Анализ сведений, имеющихся в
научной литературе, позволяет прийти к выводу, что ни одна из существующих
гипотез происхождения позвоночных не может считаться убедительной и
общепринятой. Все они малодоказательны и противоречат друг другу. Нет
убедительных сведений о происхождении симметрии, сегментации, хорды и трубчатой нервной системы.
Собственная гипотеза
происхождения позвоночных
Происхождение древних
предков человека, как и многих других животных, начинается с образования
многоклеточности. Согласно нашей гипотезе дальнейший путь таков:
создание организма типа двухслойной чаши (гастрея Геккеля Э.);
усовершенствование и усложнение этого
организма (супергастрея); колонии из супергастрей, а затем и образование многосегментного организма
(сегментелла); колонии из двух сегментелл, а затем и образование усложненного целостного многосегментного организма
(суперсегментелла); предшественники позвоночных. Мы не ставим себе целью
описать генеалогическое дерево животного мира, так как это сделать невозможно
принципиально. Речь может идти не о дереве, а о тропическом лесе, так как
происхождение животного мира полифилетическое. Наша цель выделить тот росток,
развитие которого привело к созданию человека. При этом указать место этого
ростка по отношению к предкам других представителей животного мира; также
кратко описать этапы развития этого ростка. На рис.1. представлена схема
развития позвоночных.
1. Многоклеточность. Возникновение многоклеточности шло
несколькими путями и в процессе эволюции происходило многократно. Сейчас
невозможно указать на конкретных protozoa, которые дали начало
многоклеточным организмам. Для хордовых, вероятно, многоклеточность
создавалась путем упорядоченного
деления репродуктивной клетки с последующей дифференцировкой новообразованных клеток [24]. Первые метазои, подобно
современным многоклеточным, представляли собой клон – потомков одной
единственной родительской клетки, являясь совокупностью клеток, по какой-то
причине не разошедшихся после многократного деления исходной клетки, но
сохранивших между собой связь, вероятно, с помощью внеклеточного матрикса. Этот
путь возникновения многоклеточности
можно проследить у Volvocales, а также у хордовых в
период деления–дробления яйца.
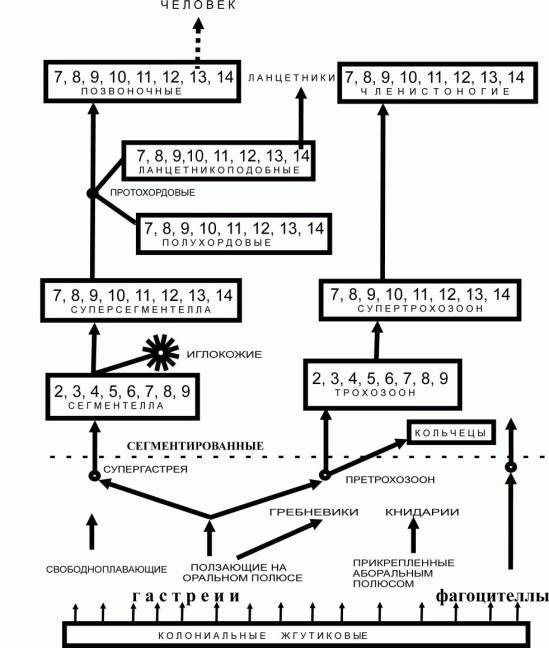
Рис. 1. Схема происхождения позвоночных
![]() - количество
агрегированных особей. Пунктирная линия отражает уровень организации животных,
выше которой отображены сегментированные животные, образовавшиеся за счет
агрегации индивидуумов. Иглокожие – агрегация сегментелл передними концами.
Членистоногие - агрегация трохозоонов передних концов с задними. Протохордовые
– агрегация сегментелл задними концами. Кольчецы – множественная агрегация претрохозоонов.
- количество
агрегированных особей. Пунктирная линия отражает уровень организации животных,
выше которой отображены сегментированные животные, образовавшиеся за счет
агрегации индивидуумов. Иглокожие – агрегация сегментелл передними концами.
Членистоногие - агрегация трохозоонов передних концов с задними. Протохордовые
– агрегация сегментелл задними концами. Кольчецы – множественная агрегация претрохозоонов.
Для колоний микроорганизмов, как и для простейших, характерно
формирование функционального органа надорганизменного уровня [18]. Наиболее
примечателен факт слияния индивидуальных наружных клеточных покровов (капсул,
экстракапсулярной слизи и др.), что ведёт к образованию единого биополимерного
матрикса [20]. В состав матрикса входят кислые полисахариды, гликозилфосфат,
биополимеры типа тейхоевых кислот, гликопротеины, также полиглутаминовая
кислота и другие биополимеры [47]. Сходство между животным и микробным
матриксами дополняется общностью некоторых химических компонентов. Матрикс
многоклеточных сообществ выполняет структурообразующую роль и относится
к надклеточному уровню организации. Благодаря матриксу колония состоит не из
одиночных клеток, а из субколониальных ассоциаций. К структуре колоний
относятся предполагаемые микроканалы для транспорта веществ - полые трубочки из
внеклеточных полисахаридов и других биополимеров. Помимо этого, через подобные
трубочки мигрируют клетки колоний [15, 52].
Влияние
внеклеточного матрикса как структурообразующего образования и нерасхождение делящихся клеток наблюдается на
разных уровнях: у первых многоклеточных, у метазоев типа гастреи Геккеля Э. и у
более сложных многоклеточных сообществ. Эти
явления выражают закон консерватизма
(фундаментальный закон эволюции материи) - природа предпочитает приспосабливать
к новообразованиям уже имеющиеся структурные и функциональные возможности,
нежели проходить путь заново или обращаться к альтернативным решениям. Эволюционный консерватизм
проявляется в том, что найденные формы
упорядочения сохраняются путем включения их в структуры следующего поколения [3].
В состоянии гомогенной массы или при наличии только
равномерного поверхностного слоя, его симметрию можно рассматривать как
симметрию шара, в котором количество осей симметрии ограничено количеством
клеток, находящихся в этом шаре. Если провести разрез через колонию в плоскости
любого меридиана, то этот разрез разделит его на две зеркальные половины, и
такая симметрия в математике называется радиальной. Фактически же такая колония
не обладала никакой биологической симметрией. В биологии симметричными
антимерами нужно признать не зеркальное отражение от какой-либо воображаемой
плоскости, а объекты, образующие сходные структуры, или объекты родственного
происхождения. Между такими частями организма может существовать полевая связь.
2. Гастрея Геккеля Э..
Разнообразие форм клеточной жизни позволило создать колонию (типа Volvox) с
признаками единого шарообразного организма, имевшего внутреннюю полость, заполненную жидкой слизью, являвшейся
внеклеточным матриксом, активно участвовавшим в регуляции множества процессов. В дальнейшем, вероятно в связи
изменением состава внеклеточного матрикса, а также из-за специализации клеток,
возможно находившихся в различных внешних условиях с двух сторон шара, а также
в связи с изменением осмотического давления вне и внутри организма, произошло
впячивание одной части шара вовнутрь с последующим формированием
пищеварительной полости с ротовым отверстием. Образовался целостный организм
наподобие примитивных кишечнополостных. Такое "изобретение" давало
ему значительное преимущество в добывании пищи и внесло революционные изменения
в развитие животного мира. В результате этого появилось множество
свободноплавающих и прикрепленных форм многоклеточных организмов.
Механизмов образования двухслойного
организма много [4, 23, 32, 46]. Возможно, у древних
форм они присутствовали и дали развитие множеству разнообразных организмов. Нам
представляется, что для источника,
который дал развитие хордовым, образование двухслойного организма наподобие
гастреи Геккеля Э. происходило в связи с влиянием кристаллоидного внеклеточного
матрикса, а также с интеграцией отдельных специализированных клеток в органоподобные
структуры. Сходную идею об интеграции разных клеток выдвинул Мечников И. И. [16] в описании развития фагоцителлы, у которой снаружи
сосредоточились клетки, имевшие реснички, а слой, обладающий фагоцитарными
свойствами и перевариванием пищи, сосредоточился внутри. Надо отметить, что у
гастреи произошла интеграция не только ресничных и фагоцитарных клеток.
Объединились и чувствительные (нервные) клетки, половые, мышечные и другие. С
этого момента колония превратилась в полноценный организм. Дальнейшее развитие
организма только усиливало интеграционные процессы. Двухслойный организм
обладал радиальной симметрией, при этом количество лучей у разных групп
животных было 2, 4, 6, 8. Источником образования супергастреии послужила
4-лучевая гастрея. Ранее уже высказывались идеи о происхождении Bilateria от четырехлучевых полипов, гастральная полость
которых была разделена на четыре камеры [55, 59, 62, 67]. Такие полипы ползали
на ротовой поверхности, их рот вытянулся и стал щелевидным, а потом сомкнулся
посередине, так, что от него осталось только два отверстия (рот и анус).
Следует добавить, что авторы этих гипотез не указывают причин, заставивших рот
вытянуться.
3. Супергастрея. Индукционные влияния от вегетативного полюса к
анимальному начинаются ещё на стадии бластулы. Подчиняясь процессам
симметрии, в соответствии с впячиванием
части организма по отношению к горизонтальной плоскости (разделяющей организм
на дорсальный и вентральный антимеры), с образованием энтодермального слоя
произошло впячивание и эктодермального
слоя. Устройство гребневиков (Ctenophora) показывает, что у них имеются
впячивания на оральном и аборальном полюсах. Первое ведет в глотку, второе
является составной частью статоциста [17]. Устройство гребневиков и
эмбриональное развитие полухордовых и хордовых показывает пути развития
полостей далеких предшественников позвоночных. Те формы первичных Eumetazoa, которые были прикреплены к субстрату
первичным ротовым отверстием, смогли образовать два отверстия в результате
неполного смыкания краев рта; через эти отверстия поступала пища, и выходили
продукты метаболизма. Дальнейший путь развития этих животных пролегал через
колонии из небольших групп особей.
Прежде чем приступить к описанию колоний, состоящих из
продвинутых гастрей, надо описать их устройство, что даст возможность понять
дальнейшие метаморфозы. Продвинутая гастрея, назовем ее супергастрея или
archeometazoa, - это не простая двухслойная чаша, а организм, прошедший сложный
путь развития. Она более сложна чем другой
гипотетический организм – галлертоид, описанный Bonik K. с соавторами [32]. Она имела
округлую форму с четким разделением на дорсальную и вентральную стороны, вела
малоподвижный образ жизни, вероятно, была ползающим животным, ориентированным
вентральной стороной к субстрату. Устройство ее несколько напоминает
гребневиков. У нее были впячивания с обеих (дорсальной и вентральной) сторон,
однако впячивания эти не были соединены между собой. Древние эуметазои имели
возможность прикрепляться к субстрату подвижным ртом, ведшим во внутреннюю
полость, которая принимала какое-то участие в пищеварении, однако, главная
функция этой полости состояла в том, чтобы быть вместилищем половых клеток,
таким образом, как это видно в развитии Сагитты (Sagitta). Продвинутая гастрея имела реснички или более
сложные структуры передвижения. Под эктодермой находилась нервная система,
состоявшая из нервных клеток, расположенных по всему телу и образующих своими
отростками сеть. Непосредственно у впячивания на аборальном полюсе было более
густое скопление нервных клеток в виде кольца. Внеклеточный матрикс (мезоглея)
сконцентрировался на аборальном полюсе
под нервным сплетением. В составе организма были железистые клетки,
вероятно выделявшие клейкие вещества.
Устройство супергастреи показано на рис. 2.
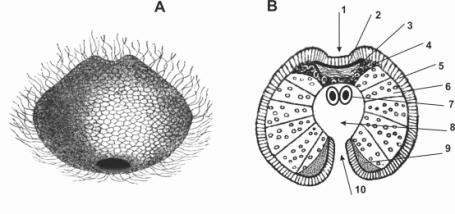
Рис. 2. Супергастрея. А – Общий вид. В – Срединный разрез.
1 – впячивание эктодермы; 2 – нервное сплетение; 3 – мезоглея;
4 – мезодерма; 5 – эктодерма; 6 – энтодерма; 7 – половые клетки;
8 – энтодермальное впячивание; 9 - железистые клетки; 10 – ротовой проход.
4. Сегментелла.
Сросшиеся своими боковыми ослизненными стенками друг с другом и соединенные
плазмодесмами отдельные зооиды объединились
в целостный сегментарный организм.
Аналогов ему в современном животном мире нет. Поэтому для удобства изложения мы
назвали этот гипотетический организм, произошедший из колоний супергастрей с
небольшим количеством особей от 2 до 9,
сегментеллой (Segmentella Yermola).
Сегментеллы с разным
количеством сегментов развивались неравномерно. Оказалось, что сегментелла с 5
сегментами была наиболее приспособлена к условиям существования и более
интенсивно развивалась. Возможно, существуют другие объяснения более широкой
распространенности 5-сегментной сегментеллы. Первоначально у сегментелл не было
разделения на переднюю и заднюю части. Однако в результате интеграционных
процессов, функционирования и адаптации к условиям жизни выделился передний
конец. Интеграционные процессы привели к образованию симметричных (относительно
плоскости, разделявшей орган на вентральную и дорсальную стороны) желобов,
которые образовались при слиянии
впячиваний, находившихся на вентральной и дорсальной сторонах
супергастрей, при этом дорсальный желоб был менее выражен. У тех сегментелл, у
которых сохранилась мезоглея, интеграционные процессы привели к сосредоточению
диффузной нервной системы вначале вокруг дорсальных впячиваний, а затем вокруг
дорсальной трубки. Таким образом формировалась трубчатая нервная система, как
это видно у полухордовых. На рис. 3 представлено поэтапное развитие описываемых
процессов.
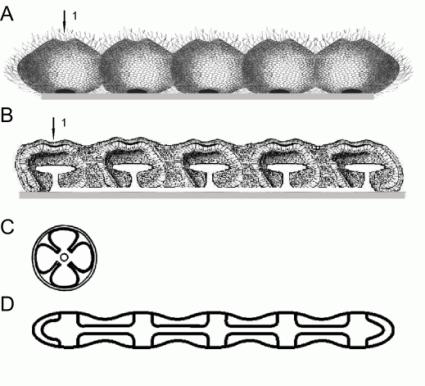
Рис.3. Образование сегментеллы.
A – колония супергастрей прикрепленная к
субстрату; B –– сегментелла, составным
элементом которой является супергастрея, на стадии образования эктодермальных впячиваний
(1 – эктодермальное впячивание); C -
строение четырех-лучевого
кораллового полипа - предка Bilateria по Ремане (Remane et al., (1989), D -
форма ротовой щели (сквозные поперечные отверстия и межсегментные ходы), вид с
оральной стороны.
С расположением особей в ряд
образовалась дополнительная плоскость симметрии (сагиттальная). При этом
сформировался организм с радиально-двухплоскостной (радиально-четырехсторонней)
симметрией. Одна плоскость симметрии разделяла организм на дорсальный и вентральный
антимеры, а вторая, сагиттальная, разделяла каждый из этих антимеров еще на 2
части. Так образовалось 4 антимера. В связи с организацией такой симметрии
количество антимеров увеличилось за счет разделения первичных сегментов на 4 части при их метамерном расположении.
В соответствии с трохофорной
гипотезой [1] кольчецы и членистоногие произошли от гипотетического предка -
трохозоона, имевшего трохоподобное строение, который в свою очередь произошел
от предков ползающих гребневиков. На наш взгляд, трохозоон соответствует по
развитию сегментелле, он также имел
сегментное строение. Различие между сегментеллой и трохозооном заключалось в
том, что у последнего интеграционные процессы отдельных зооидов в целостный
организм были более выражены. Кроме того, мезоглея предков исчезла, а мезодерма
была более выражена.
5. Суперсегментелла.
Сегментеллы развивались как отдельные организмы, а также объединялись в
колонии. Одновременно существовали как свободные сегментеллы, так и
колониальные. Сегментеллы с разным количеством сегментов создавали колонии,
однако наиболее жизнеспособными оказались колонии, где 5-сегментные сегментеллы
объединялись с другими сегментеллами, имевшими разное количество сегментов (от
2 до 9). Следовательно, в новом организме было от 7 до 14 первичных сегментов.
Надо отметить, что несколько сегментелл, объединившихся передними концами
радиально (вокруг общего рта), стали предшественниками иглокожих. В других
группах объединение двух сегментелл, состоящих из разного количества особей
(первичных сегментов), в единый организм шло путем соединения задних концов что
привело к усложнению общей организации тела. Такой организм мы назвали
суперсегментелла.
Сходные процессы происходили
в развитии трохозоона, где агрегация множества претрохозоонов в ряд дала
развитие кольчецам (annelides). А двухкомпонентные организмы, имевшие в своем
составе от 7 до 14 сегментов (по формуле 5+n; где n = от 2
до 9) дали развитие всем членистоногим (arthropoda), при этом соединение двух
разносегментных контактирующих организмов происходило передним концом к
заднему.
Можно предполагать, что
суперсегментеллы были малоподвижны или были прикреплены к субстрату.
Способность объединятся головными и каудальными концами отдельных сегментированных организмов
сохранилась у некоторых современных видов аннелид [1]. В результате того, что
после деления особи не расходятся, образуется колония, которая в таком виде
может существовать длительное время.
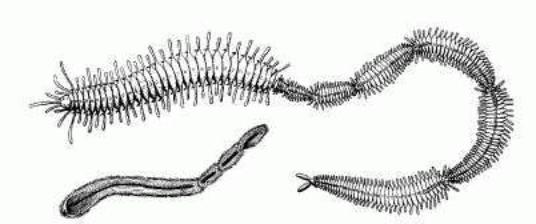
Рис.4. Временная колония червей по Н.К. Янковскому (2007)
На рис. 4 представлена
линейная колония червей по Янковскому Н. К. [26] Линейные колонии сегментелл
постепенно превратились в целостные организмы, как это неоднократно было ранее
в развитии многоклеточных, когда колония превращается в целостный организм. С
присоединением к 5-сегментному организму другого сходного организма создавалась
ситуация, когда общее устройство отдельных соединяемых организмов имело
радиально-двухплоскостную симметрию, а количество антимеров соответствовало
радиально-трехплоскостной симметрии. В связи с этим произошла реорганизация
всего организма. У
суперсегментеллы произошло объединение
дорсальной и вентральной трубок с образованием U-образной полости,
принимавшей участие в пищеварении. U-образный кишечник
образовался и у трохозоона. У них, как и у предков крыложаберных, мезоглея не оказывала влияния на формирование
дорсальной трубки. У этих животных образовался U-образный кишечник, но в нем
не было концентрации нервной ткани. Короткое тело отдельных зооидов Rhabdopleura, относящихся к крыложаберным (Pterobranchia), как бы сложено вдвое, так
что анальное отверстие открывается на переднем конце позади головы. Как
известно, эта группа животных имеет очень слабое сходство с хордовыми, так как
у них нет полой нервной трубки и хорды. У некоторых суперсегментелл ещё не
завершился процесс формирования U-образной трубки, как
произошло обособление дорсальной трубки вместе с загибом. Эта часть трубки
выпрямилась и обе структуры стали самостоятельными, приобрели новые функции.
Дорсальная трубка стала выполнять функцию центральной нервной системы, а
вентральная - пищеварительную. Интеграционные процессы привели к образованию
нового организма. Края впячиваний всех
сегментов сомкнулись на дорсальной и вентральной сторонах, при этом первичная пищеварительная
полость превратилась в кишку с новыми собственными ротовым и анальным отверстиями. Межсегментная ротовая щель превратилась в эндостиль, а первичные
отверстия, образовавшиеся из-за несмыкания рта, - в жаберные щели. Новый
организм получил возможность свободно передвигаться. Но, будучи фильтратором по
образу жизни, суперсегментелла эту возможность вначале не использовала. Так сформировались
организмы наподобие ланцетника.
6. Предшественники
позвоночных. У других суперсегментелл процесс интеграции нервной
системы продолжался, и в ответ на
образование загиба в задней части, в соответствии с симметрией (по отношению к
плоскости, разделявшей переднюю и заднюю части тела) образовался загиб в
передней [4, 13] (см. рис.5).
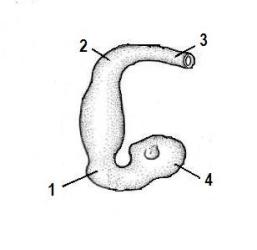
Рис.5. Ранняя стадия развития головного мозга человека по С. Гильберту (1993).
1 – головной изгиб; 2 – шейный изгиб; 3 – спинной мозг 4 – передний мозговой пузырь
Однако полного объединения
дорсальной и вентральной трубок в переднем отделе не произошло. Последовало
выпрямление загибов дорсальной трубки с образованием трубчатой нервной системы
и кишечника. При этом в результате интеграционных процессов увеличилось
количество нервной ткани как на переднем, так и на заднем концах, которые стали
предшественниками головного мозга и мозжечка. Напоминанием того, что нервный
канал и пищеварительная полость были единым целым, служит наличие
нервно-кишечного канала у всех позвоночных.
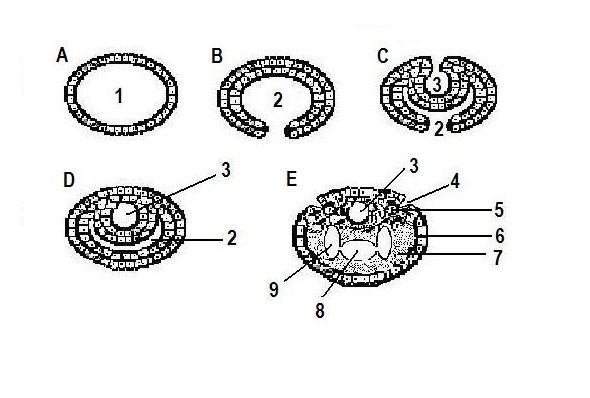
Рис. 6. Схема развития полостей гипотетического предкового организма - предшественника позвоночных
A – E – этапы развития полостей в соответствии с симметрией.
1 – первичная полость; 2 - энтодермальное впячивание; 3 - эктодермальное впячивание (нервная трубка); 4 – полость боковой линии; 5 – мезодерма; 6 эктодерма; 7 – энтодерма;
8 – полость первичной кишки; 9 – целом.
На рис. 6 представлена схема
развития полостей организма (первичной кишки и нервного канала) как антимеров,
а затем нервной трубки и полости боковой линии, первичной кишки и целома. В
связи с различными функциями первоначально симметричных образований (пищеварительной
полости и нервной трубки; пищеварительной полости и целома), а также вначале
функционально связанных образований (пищеварительной полости и входных-выходных
отверстий) шли морфологические изменения, приведшие к потере родственных
отношений этих структур и развитию их как самостоятельных образований. В
филогенетическом развитии первичная пищеварительная полость преобразовалась в
кишечник и целом. Об этом говорит энтероцельный способ образования целома у
всех хордовых. Первичный же целом дает начало всем полостям тела взрослого
организма [12]. Первичные входные-выходные отверстия, сосредоточившись в
передней части тела, стали жаберными отверстиями. В последующем в связи с
приобретением ими функции "дыхания" и развитием вокруг них
рецепторов, а также в связи с общими процессами интеграции первичные
пищеварительные отверстия изменили свою морфологию и место расположения.
Обобщая изложенное в данной статье, надо подчеркнуть,
что новые органы не появились из ничего. Имеющиеся у многоклеточных организмов
ткани и клетки адаптировались к меняющимся условиям, специализировались,
располагались в организме в соответствии с симметрией, присущей ему на данный
период развития. Мезоглея, пройдя сложный путь развития, являлась организатором
первичных многоклеточных, в конечном итоге стала хордой, следовательно, стала
организатором хордовых животных. Из нервных сплетений, имеющихся на аборальном
полюсе, образовалась узловая нервная система, из диффузной нервной системы,
которая концентрировалась вокруг эктодермальных впячиваний, а затем
эктодермальной трубки, произошла трубчатая нервная система. Из железистых
клеток образовалась железистая ткань в каждом сегменте.
С окончания
действия хорды как организатора развившаяся мезодерма стала организатором в построении позвоночных.
Первая плоскость симметрии у древних многоклеточных разделяла организм на
дорсальную и вентральную стороны. С расположением сегментов в ряд организм
приобрел сагиттальную плоскость
симметрии, разделявшую организм на правую и левую стороны (при определившемся
переднем конце тела). При комбинации двух организмов появилась третья плоскость
симметрии. Организм длительное время был организован как трехплоскостная
система. Однако, в связи с функцией органов родственные связи органов,
находящихся по разные стороны
плоскости, ранее разделявшей организм на дорсальную и вентральную
стороны, были утеряны. Следовательно была исчезла одна плоскость симметрии. И
организм стал симметричен по двум сторонам (сагиттальной и относительно
плоскости, разделяющей организм на передний и задний концы). Изменения в структуре организма относительно
сагиттальной плоскости происходили незначительные, поэтому билатеральная
симметрия четко выражена, однако без особого труда находятся антимеры двух и
даже трехплоскостной симметрии. Не надо забывать, что при образовании гастреи
первичной была радиальная симметрия, и эта симметрия присутствует в организме
человека. Так
образовались первичные хордовые, предшественники позвоночных.
Заключение
Предложенная нами гипотеза
происхождения позвоночных затрагивает важнейшие вопросы развития Eumetazoa.
Мы не планируем в данной работе представить осветить все вопросы
формообразования, а решили высказать свою точку зрения о наиболее важных
аспектах этой проблемы.
1. Филогения
На наш взгляд общей ошибкой большинства
исследователей, кто пытается решить вопрос происхождения хордовых, является
стремление найти «достойного»
представителя беспозвоночных и интрополировать какие-то его свойства на
древнего предка, который мог развиваться несколько иначе и дать начало всем
хордовым. Мы думаем, что такой подход не верен. Дело в том, что в
археологической летописи до сих пор нет сведений об этих древних формах жизни.
Реконструировать древнего предка позвоночных и пути его развития можно, только
исходя из общих тенденций развития всех животных. Такую попытку мы сделали в
изложенном материале этой статьи.
Сходство в строении тела анелид и членистоногих с
позвоночными несомненно, если сравнивать их строение в зеркальном изображении.
А отсюда родились идеи их общего предка, в частности urbilateria и
перевернутости брюшной и спинной сторон рассматриваемых животных. На наш
взгляд такой предок был, и это подтверждается молекулярной биологией, только
вопрос состоит в том, где его место в филогенезе, и что привело к
противоположному расположению органов у этих групп животных. По нашему
представлению, общим предком этих животных была несегментированная
супергастрея, имевшая радиальную четырех-лучевую симметрию. Гипотетическое
животное urbilateria (общий сегментированный
предок всех bilateria по De Robertis Е. (36) вряд ли
когда-либо существовало, а сходство в строении предков позвоночных и
трохофорных животных объясняется тем, что в основе колоний находился один и тот
же организм (гастрея) с одинаковым способом организации осевой симметрии
(агрегация индивидуумов в ряд в колонии, с последующим преобразованием ее в
целостный организм). Различия двух ветвей дивергировавших родственных
организмов объясняется различием и местоположением в организме организатора.
Для трохофорных животных это мезодерма, а у предков хордовых (сегментелла Yermola) это, вначале
мезоглея, а потом мезодерма.
2. Функция
Важной ошибкой некоторых исследователей является
попытка объяснить появление главных
структур позвоночных, их симметрию и сегментацию функцией организма, или одного из органов, средой обитания,
образом жизни, формой движения. Эти факторы и связанная с ними адаптация
являются механизмами эволюции и вносят существенный вклад в трансформацию организма,
однако, существуют другие механизмы эволюции, на которые они никак не влияют.
При этом надо обратить внимание, что функция может изменить существующий орган,
но она вовсе не способна создать какой-либо новый орган или организовать
симметрию. Функция организма связана с внешней средой, которая может только
уменьшить количество плоскостей симметрии. В соответствии с принципом Кюри П.
[35] у объекта остается только та симметрия, которая совпадает с симметрией
среды. Симметрию организма определяет ее внутренний состав, имеющий
кристаллоидное строение [40]. Часто образование новых органов объясняют
изменившимися условиями существования, и появлением в связи с этим потребности
организма в тех или иных органах. Объяснения такие некорректны. Если бы такие
механизмы существовали, то жизнь не развивалась бы выше простейших, которые
существуют более 3,3 млрд. лет и значительно не изменились. Объяснение
появления органов за счет потребности в них организма некорректно, так как создание чего-либо в вязи с потребностью является проявлением воли, а это связано с
креатором.
3. Интеграция
Некоторые авторы публикаций на биологические темы
объясняют интеграционные процессы функцией организма и средой обитания. На
самом деле, интеграционный процесс – выражение глобального закона объединения
родственных клеток. В эволюции проявлением этого закона явилось образование и
функционирование половых клеток. В дальнейшем путем объединения родственных
клеток (одноклеточных организмов) создавались колонии и целостные организмы.
Интеграция и специализация являются эволюционными механизмами. В многоклеточном
организме специализированные клетки объединялись, образуя из имеющегося
материала новые органы. Глобальный закон интеграции выражается в объединении
отдельных многоклеточных индивидуумов в
колонии, а затем в целостный организм. Этот же закон действует и на более
сложных уровнях организации живого.
4. Организаторы
Жизнь зарождалась путем
кристаллизации и полимеризации органических и неорганических веществ. Сама
клетка образовалась на основе кристаллических или кристаллоподобных
организаторов. Слизеподобная мезоглея первых шарообразных организмов и явилась
внеклеточным организатором. Имеющийся у
всех многоклеточных животных внеклеточный матрикс (ВКМ) активно участвует в
регуляции множества процессов. Он определяет форму клеток, их миграцию,
пролиферацию и дифференцировку. Мезоглея у кишечнополостных выполняет функцию
скелета [31, 34, 65]. У некоторых представителей кишечнополостных мезоглея
приобретает сходство с соединительной тканью [7, 15, 33]. С развитием жизни состав внеклеточного
матрикса изменялся. Изменялись его свойства и функция. У полухордовых внеклеточный матрикс выглядит в виде
небольшого тяжа студенистого вещества, окруженного одним слоем клеток [19, 56,
58]. Тем не менее, эта структура, называемая хордой, является организатором,
что отражается в раннем эмбриогенезе в развитии хордовых, в том числе и
позвоночных.
На самых ранних стадиях
развития у двухслойных организмов типа гастреи Геккеля Э. появляется слой
клеток мезодермы, которая постепенно приобретает ведущую роль в организации
организма. Уже у сегментеллы она является главным индуктором интеграционных
процессов, она является организатором организма. Существует логичное мнение
[14], что первичный рот (бластопор), являясь великим организатором, разделился
на две части (ротовое и анальное отверстие) и таким образом мог разделить единый организатор надвое.
Однако рот, как организатор, строит структуры радиальной симметрии.
Ланцетникоподобные организмы и позвоночные имеют два организатора с признаками
билатеральной симметрии, поэтому логичнее предположить, что такая ситуация
сложилась в результате объединения двух самостоятельных организмов, имеющих
плоскостную симметрию.
5. Симметрия
Беклемишев В. Н. [1], Северцов А. Н. [21]
предполагали, что симметрия тела всех Bilateria
гомологична. Как видно из этого названия, симметрия этих животных двухсторонне
симметрична. При этом подразумевают единственную сагиттальную плоскость. Как мы
уже говорили в разделе изложения своей гипотезы симметрия этих организмов
значительно сложнее. Что касается симметрии относительно сагиттальной
плоскости, гомологичность ее полностью (у всех животных) не может быть
достоверной. Однако у всех сегментированных животных, у которых сегменты
расположены в ряд, такая симметрия гомологична (механизм ее образования
идентичен). Но существуют несегментированные животные с признаками
билатеральной симметрии, которая не может быть признана гомологией таковой
сегментированных животных. Примером могут служить щетинкочелюстные (Caetognatha), в частности виды рода Сагитты (Sagitta), обладающая двусторонней симметрией. Исходно предки
этого животного обладали радиально-двухлучевой симметрией. Продольное
вытягивание тела позволяет рассматривать это животное как обладающее
радиально-двухсторонней симметрией.
6. Гомеобоксные гены
Сравнительный анализ структуры и экспрессии
гомеобоксных генов позволят изучать сегментацию и гомологию органов, они регулируют развитие основных структур,
сегментацию, закладку конечностей и другие процессы. Считается, что эти гены
возникли в результате множественной дупликации одного предкового гена [5, 14].
Такие гены называются паралогами. На основании такого представления о
происхождении кластера гомеобоксных генов строятся гипотезы о происхождении
сегментации и двух организаторов, и о постепенном наращивании количества
паралогов и, следовательно, сегментов. Известно, что у позвоночных максимальное
число паралогов кластера гомеобоксных генов составляет 13, а у ланцетника - 14
[42]. На основании этого Городилов Ю. Н. [5] делает вывод, что у ланцетников в
процессе эволюции за период свыше 500 млн. лет в пределах цепочки одного
комплекса Нох не только сохранились все гены этой группы, но их число
выросло.
Альтернативой этим мнениям может быть предположение об
объединении генов индивидуумов в
результате объединения их самих сначала в колонию, а затем в единый организм.
При этом надо учесть, что объединение самостоятельных организмов происходило в
двух группах, а потом произошло объединение этих групп. Количество сегментов
(индивидуумов, генов) в группах выражается в формуле 5 + n, где n равно от 2 до
9. Общее количество сегментов составляет от 7 до 14. Такая трактовка
происхождения так называемых паралогов позволяет объяснить разное их количество
у позвоночных и хордовых, а также происхождение двух организаторов. Есть
сведения, что организаторы Шпемана - головной и туловищный - являются двумя
отдельными, совершенно независимыми структурами, имеющими разное происхождение
[5]. Подтверждением этому служат опыты, которые показывают, что при инактивации
генов Liml, Otx2 приводит к
полному отсутствию головы, при этом туловищно-хвостовая часть тела развивается
нормально [5, 27].
Малахов В. В. [14] в обзоре литературы о происхождении
хордовых разбирает гипотезу Дьюел Р. [37] о происхождении хордовых путем
превращения колоний особей кишечнополостных в единый организм и он указывает,
что в эмбриогенезе Bilateria нет никаких следов
слияния нескольких особей. Однако факт этот не противоречит вероятности
согласованного действия генов отдельных особей, находящихся в единой структуре.
Известно о согласованном действии генов в опытах при получении аллофенных мышей
[4].
7. Рот и его функции
Образование рта явилось великим прорывом в развитии
животного мира. С его образованием изменилась форма пищеварения, что в
дальнейшем привело к значительной реорганизации организма. Однако роль рта не
сводится только к функции некоего клапана, регулирующего поступление пищи и
выведение продуктов метаболизма. Его главная роль заключается в организации
организма. Впячивание у бластеи привело к образованию рта и шарообразной
пищеварительной полости. Многие авторы, моделировавшие события, приведшие к
преобразованию этих структур в кишечник с входным и выходным отверстиями, не
предоставили убедительных доказательств правильности их моделей. Длительное
время считалось, что организм типа гастреи по каким-то причинам вытянулся в
длину, а, следовательно, вытянулась и пищеварительная полость, и в связи с
необходимостью выведения продуктов обмена веществ из длинной пищеварительной
полости образовалось второе (выходное) отверстие. А потом по каким-то причинам
это выходное отверстие стало входным, таким образом, поменялась функция
отверстий и направленность движения пищи по пищеварительному тракту. На этих
основаниях биологи делили животных на первично- и вторичноротых. Малахов В. В.
[14], как и другие современные исследователи, изучавшие происхождение
билатеральных животных, проанализировав сведения по филогении животных,
методами сравнительной анатомии, сравнительной эмбриологии, палеонтологии и
молекулярной биологии, пришли к выводу, что рот и анус произошли от переднего и заднего концов удлиненного
бластопора. Они подтверждают гомологию брюшной стороны Bilateria и орального диска кишечнополостных. Однако он ничего
не говорит о причинах, заставивших бластопор вытянуться, а потом сомкнуться
посередине с образованием рта и ануса. С принятием такой трактовки образования
рта и ануса отпадает вопрос о первично- и вторичноротости животных, тем более что реальная
первичноротость характерна только для плоских червей, а истинная
вторичноротость свойственна иглокожим [19].
Формирование билатеральной
симметрии начинается у билатерий только после стадии гаструлы. Бластопор
вытягивается и приобретает щелевидную форму, потом бластопор замыкается с образованием двух отверстий (рот и анус).
И такая последовательность событий эмбриогенеза билатерий легко объясняется
нашей гипотезой. Вначале односегментный организм из бластеи превратился в
гастрею, а потом последовательная агрегация гастрей в ряд обеспечила симметрию
относительно саггитальной плоскости. Соединение ротовых отверстий нескольких
особей дало удлиненный рот. Края рта, имея сродство, интегрировали в единую
структуру с образованием пищеварительной полости со ртом и анусом.
1.
Происхождение
трубчатой нервной системы
Ранее высказывалась идея
создания трубчатой нервной системы путем скопления нервных клеток вокруг
пищеварительного канала у паукообразных (цитировано по Савельеву С. В. [19].
Эта идея была подвергнута жёсткой критике в связи с тем, что для ее принятия
нужно сделать ряд невероятных предположений о развитии нового кишечника и
хорды. Эти преобразования не отражаются в эмбриональном развитии позвоночных.
Однако такая идея в обобщенном виде заслуживает внимания, так как нацеливает
исследователя на понимание механизма образования нервной системы путем
скопления нервных клеток вокруг готовой полости (трубки). Новая гипотеза
Савельева С. В. [19] о создании нервной трубки путем интеграции нервных стволов
в единую структуру вносит новое в понимание вопроса, он предполагает, что
полость образовалась в связи с потребностью нервных структур в циркуляции
жидкости вокруг нервных клеток для улучшения метаболизма. Во-первых, как мы уже
говорили, потребностью нельзя обосновать образование каких-либо органов или
структур. Во-вторых, автор не упоминает, какова функция этой жидкости у древних
предков хордовых.
На основании анализа
сведений, имеющихся в научной литературе, мы пришли к выводу, что фактически
нет обоснованных идей образования нервной трубки, а, следовательно, нет
обоснованных идей происхождения хордовых.
Наша гипотеза образования нервной трубки как слившихся аборальных
впячиваний сегментированного организма показывает пути образования хордовых.
ЛИТЕРАТУРА
1. Беклемишев
В. Н. Основы сравнительной анатомии беспозвоночных. // «Наука», Москва, 1964.
2.
Воронов
Д. А.
Старая гипотеза «перевернутости» хордовых подтверждается. Природа (2000)
11, рр. 18-22. http://learnbiology.narod.ru/23.htm
3. Галимов Э. М. Феномен жизни. Между равновесием и нелинейностью; происхождение
и принципы эволюции. // Москва, 2001.
4.
Гильберт
С. Биология развития. // «Мир», Москва, 1993.
5.
Городилов Ю. Н. Исследования раннего онтогенеза в
связи с проблемами филогении и происхождения позвоночных животных. // Дис. ...
д-ра биол. наук: Санкт-Петербург, 2002.
6.
Дорн
А. Происхождение позвоночных животных и принцип смены функций. //
«Биомедгиз», Москва –Ленинград, 1937.
7.
Заварзин
А. А. Очерки эволюционной гистологии крови и соединительной ткани. // «Медгиз»,
Москва - Ленинград, 1953: том 4.
8. Иванов А. В.
Современная эволюционная морфология. // «Наука», Москва, 1991.
9.
Иванов
А. В., Мамкаев Ю. В., Ресничные черви (Turbellaria), их происхождение и
эволюция. Филогенетические очерки. // «Наука», Ленинград, 1973.
10.
Иорданский
Н. Н. Эволюция жизни. // «Академия»,
Москва, 2001.
11.
Карлсон Б. Основы эмбриологии по Пэттену. // «Мир», Москва, 1983:
том 1, стр. 173- 179.
12.
Карлсон
Б. Основы эмбриологии по Пэттену. // «Мир», Москва, 1983: том 1. стр. 35.
13.
Коуэн
У. Развитие мозга. В кн: Мозг. «Мир», Москва, 1984: стр.116.
14.
Малахов
В. В. Происхождение билатерально-симметричных животных (Bilateria).
// Журнал общей биологии (2004) 65, №5, стр. 371-388
15. Матвеев И. В. Экспесссия
гена мезоглеина в разных типах клеток медуз Aurelia aurita.
// Институт цитологии РАН, . Дисс. Канд. Биол. Наук, Санкт Петербург, 2007.
16. Мечников И. И.
Сравнительно-эмбриологические исследования. III. О гаструле некторых многоклеточных. // В кн.: Академическое
собрание сочинений. «Госиздат мед. лит.», Москва, 1955: том 3, стр. 104-24.
17.
Натали
В. Ф. Зоология беспозвоночных. // «Государственное учебно-педагогическое
издательство», Москва, 1951: стр. 148.
18. Олескин А.В.
Надорганизменный уровень взаимодействия в микробных популяциях. // Микробиология
(1993) 62. № 3, стр..389-403.
19.
Савельев
С. В. Происхождение мозга. // «Веди», Москва, 2005.
20.
Сафронова
И. Ю., Ботвинко И. В. Межклеточный матрикс Bacillus subtilis 271:
полимерный состав и функция. // Микробиология (1998). 67, № 1,
стр. 45.
21. Северцов А. Н. Собрание сочинений. // «Изд-во АН
СССР», Москва-Ленинград, 1945: том 3.
22. Сент-Илер Э. Ж. О позвонке у
насекомых // Избр. тр. / Ред. И.Е.Амлинский. Москва, 1970: стр. 375 - 390.
23. Серавин Л. Н., Гудков А. В. Trichoplax Adhaerens (тип Placozooa) одно из самых примитивных многоклеточных животных. // Учебное пособие.
Санкт-Петербургский Государственный Университет, Институт цитологии РАН. Санкт-Петербург, 2005.
24. Фролов Ю. П. Некоторые аспекты
происхождения Metazoa. // Вестник СамГУ – Естественнонаучная серия.
(2006). №7 (47), стр. 241.
25. Юшкин Н. П. Биоминеральные
взаимодействия М.»Наука»2002
26. Янковский Н. К. Лекции по
биологии. // http://bio.fizteh.ru/student/files/biology/biolections/
27. Acampora D., Masau S., Lallemand Y. Avantoggiato V.,
Mauri M., Simeone A. and Brulet P. Forebrain and midbrain regions arc deleted
in Otx2-/- mutants due to a defective anterior neuroectoderm
specification during gastrulation // Development (1995) 121, pp.
3279-3290.
28. Arendt
D. and Nübler-Jung K. Comparison of early nerve cord development in insects and vertebrates. // Development (1999) 126, pp.
2309-2325.
29. Arendt
D., Nubler-Jung K. Inversion in dorsoventral axis? // Nature. (1994). 371,
pp. 26;
30. Berrill N.J. The Origin of Vertebrates. // Oxford
Univ. Press, London, 1955.
31. Bonillon Y. Coppois G. Efude
comparative de la mesoglee des cnidaires. // Cahier Biol. Marine (1977) 13,
pp. 339-368.
32. Bonik K., Grasshoff M., Gutmann W.F. 1976. Die
Evolution der Tierkonstruktion. // Natur und Museum 106, pp.
129-143.
33. Chapman
D.M. Cnidarian histology. Coelenterate biology. Reviews and new perspectives.
Eds. Muskatine L., Lenho H. Acad. Press. NY - SF - London. 1974: pp. 1-93.
34. Chapman G. 1966. The structure
and function of the mesoglea. Cnidaria and their evolution. // ed. Rees. Symp.
Soc. London: 16 pp. 147-168.
35. Curie P. Oeuvres.// Preface, Paris, 1908.
36. De
Robertis E.M. and Sasai Y.. A common plan for
dorsoventral patterning in Bilateria. // Nature (1996) 380, N 7, pp. 37
- 40.
37. Dewel
R.A., Colonial origin for Eumetazoa: major morphological transitions and
the origin of bilaterian complexity // J. Morph. (2000) 243, pp.
35-74.
38. Ermolenko A.E. The General Plan of Human Structural Organization. //
Bibliographical Index of VINITY
"Deposited Scientific Works" 1996; № 12; n 121. Deposited at VINITI, N 3105 B-96. Moscow
1996.
39. Ermolenko A & Perepada E Origin of segmentation in the human
structure. // Medical Hypotheses (2006)
67, pp. 622-625.
40. Ermolenko A. E., Perepada E. A. The biocrystalloid structure of man:
an extracellular theory Acta Bio Medica. Vol. 78 – suphl.
1/2007 pp. 21-25.
41. Erwin D.H. The Origin of Bodyplans. // Amer. Zool.,
(1999) 39, pp. 617-629
42. Foreman R.E., Gorbman A., Dodd J.M., Olsson R. Evolutionary
Biology of Primitive Fishes. //: Plenum Press, N.Y. 1985: Vol. 103. - NATO ASI
Series A.
43. Garsia-Fernandez
J., Ferrier D.E.K., Minguillon C, Holland P.W.H. and Wada H. The Amphioxus
genome has both archetypal and derived features // Zool.Sci. (2001) 18,
pp. 454-455.
44. Gerhart
J., Lowe C, , Kirschner M. Hemichordates and the origin of chordates. // Curr Opin Genet Dev. (2005) 15, N 4, pp. 461-467.
45. Gerhart J The deuterostome
ancestor. //J Cell Physiol. (2006).
209, N 3, pp.677-85.
46. Grasshoff M. 1993. Die Evolution der Tiere in neuer
Darstel-lung. // Natur und Museum. 123, pp. 204-218.
47. Gygi
D., Rahmen M.M., Lai H.-C., Carlson R., Guard-Petter J., Hughes C. A cell
surface polysaccharide that facilitates rapid population migration by
differentiated swarm cells of Proteus mirabilis // Mol. Microbiol.
(1995) 17. pp.1167-1175.
48. Hyman
L.H., The Invertebrates. Plamyhel-minthes and Rhynchocoela. The acoelomate
Bilateria. // N. Y.: McGraw-Hill Book Co, 1951.
49. Jagersten
G.,. On the early phylogeny of the Metazoa. The Bilaterogastraea-theory //Zool.
Bidr. Uppsala. (1955) 30. pp. 321-354.
50. Jefferies R.P.S. The Ancestry of Vertebrates. //
Lond.: British Museum, 1986.
51. Jollie M. What are the «Calcichordata»? and the Larger
Question of the Origin of Chordates // Zool. J. Linn. Soc. (1982), 75,
pp. 167-188.
52. Kleinman
HK, Philp D, Hox man MP. Role of the extracellular matrix in
morphogenesis. // Curr Opin Biotechnol, (2003) 14 pp. 526-532.
53. Kuhlenbeck H. Invertebrates and Origin of Vertebrates. // Karger Press,
Geneva, 1967.
54. Lipps J.H., Signor W. Origin and Early Evolution of the Metazoa. //
Plenum Press, N.Y., 1992.
55. Marcus E. On the evolution of the animal phyla //(1958) Quart. Rev.
Biol, 33, pp 24-58.
56. Meglitsch P.A., Schram F.R. Invertebrate Zoology. // Lond.:
Oxford Univ. Press, 1991.
57. Nielsen C. Origin of the chordate central nervous system - and the
origin of chordates. Development Genes and Evolution, (1999) 209, N 3, pp. 198-205.
58. Olson R., Holmberg K., Lilliemarc Y. Fine Structure of the Brain and
Brain Nerves of Oikopleura dioica (Urochordata, Appendicularia) // Zoomorphology.
(1990). 110, pp. 1-7.
59. Remane A., Starch V., Welsch U., Kurzes Lehrbuch der Zoologie. //
Gustav Fischer, Stuttgart, 1989: pp. 572.
60. Rieger R.M., Evolution of the "lower" Metazoa // Early life on
earth. Nobel Symposium 84 / Ed. Bengtson S. N.Y.: Columbia Univ. Press. 1994,
pp. 475-488.
61. Slack J.M.W., Holland P.W.H., Graham C.F. The zootype and phyloptypic
stage. // Nature (1993)
361, pp. 490-492.
62. Slewing R., Das Archicoelomatenkoncept // Zool. Jb. Abt. Anat, (1980).103,
pp. 439-482.
63. Spemann H. Embryonic Development and Induction. //
Reprinted 1962 by Hafner Publishig Company, New York, 1938: pp. 401.
64. Spemann GG Mangold H. Ueber Induction von Embryonalanlagen durch
Implantation ortfremder Organistoren.// Arch. microsk. Anat. Entwmech,
(1924) 100, pp. 599-638.
65. Weber C., Schmid V.
The fibrous system in the extracellulare matrix of hydromedusae. // Tissue
and Cell. (1985) 17, pp. 811-822.
66. Willmer P. 1994. Invertebrate relationships: pattern
in animal evolution. Cambridge: Cambridge Univ. Press. 397p.
67. Ulrich W.
Vorschlaege zu einer Revision der Grosseinteilung des Tierreichs // Verh.
Deutsch. Zool. Ges. Marburg. 1951: pp.
215-271.
Origin of vertebrates
Ermolenko
A., Perepada E.
We suggest a new hypothesis of the origin of vertebrates.
Development of vertebrata
begins with formation of a multicellular organism by ordered repeated division of a reproductive cell and
nondisjunction of the new formed cells, which have kept connection by means of the
extracellular matrix.
Further there is a consecutive formation of organisms due to aggregation
of similar structures. In such a way were formed: Gekkel’s blastaea; Gekkel’s
gastraea; advanced gastraea (supergastraea), an organism like creeping
ctenophore; Yermol’s segmentella which consisted of a small amount (2-9) of
supergastraeas, an organism like oligomeric worms; supersegmentella formed by
connection of posterior edges of two
segmentellas. Supersegmentellas gave development to tunicates, hemichordates,
chordates like lancelet and to predecessors of vertebrata.
Origin of organs: from extracellular matrix of the first multicellular
organisms originated mesogloea, and then a chord; endodermic embolies gave
development to the primary gut; ectodermic embolies after the concentration
there earlier diffusely located nervous cells transformed first into a trench,
and then into a tubular nervous system; the condensed nervous fabric of aboral
poles gave development to the central nervous system. Glandular cells of
supergastraea were the derivative material for all glands according to the primary
segmentation.
Segmentation of
organisms is determined by aggregation of supergastraeas into one integrated
organism with two-plane symmetry.
The origin of symmetry: symmetry is determined by structure-forming
quality of extracellular matrix. Symmetry of primary organisms was radial; then
radial- bilateral, and the first plane of symmetry divided the organism into
dorsal and ventral sides. With the arrangement of supergastraeas in a line
radial-two-plane symmetry consistently formed, thus the new sagittal plane
divided the organism into right and left antimeres. Radial-three-plane symmetry
formed by association of two organisms (segmentellas) by posterior edges.
The third plane of symmetry divided the organism into anterior and posterior
antimeres.
Key words: chordates, vertebrates, symmetry, nervous
system, gastraea, extracellular matrix, primary gut.
* Обзор литературы и резюме были опубликованы ранее: Математическая морфология. Т. 6. Вып. 2. - 2007. Здесь представлена собственная гипотеза, а для удобства пользования материалом представлен и ранее опубликованный обзор литературы.
ФГУ НИИ трансплантологии и
искусственных органов Росмедтехнологий
Поступила в редакцию
22.01.2008.