ЭКСПЕРИМЕНТАЛЬНАЯ
МОРФОЛОГИЯ . XXI ВЕК
УДК
611.13/.16+611.423]:616-006-092.9
КЛЕТОЧНАЯ И ТКАНЕВАЯ ИНЖЕНЕРИЯ ЭНДОТЕЛИЯ
ФОРМИРОВАНИЕ В КУЛЬТУРЕ ЭНДОТЕЛИЯ IN VITRO
ФУНКЦИОНИРУЮЩИХ САМОРАЗВИВАЮЩИХСЯ КАПИЛЛЯРНЫХ СЕТЕЙ.
ЭКСПЕРИМЕНТАЛЬНЫЕ ПОДХОДЫ
© 1997 г. В. А. Глотов
Разработана
концепция реактора для запуска в
культуре эндотелия in vitro процесса
формирования функционирующих
саморазвивающихся капиллярных
сетей, базирующаяся на современных
достижениях микроэлектроники в
области больших интегральных
микросхем. Разработаны эскизы
технических заданий для НИР и ОКР
такого реактора. Систематизированы
формообразовательные феномены в
культуре эндотелия in vitro, собранны
данные по их прикладной инженерной
морфометрии. Начаты НИР и ОКР по
отдельным системам реактора.
Определены условия, в том числе и
финансовые, начала совместных
работ со специализированными
предприятиями. Проведены работы по
биофизико- и
математико-морфологическому
моделированию морфологических
феноменов в микрососудистых сетях,
необходимые для разработки
гибридной интегральной
электронной биогидромеханической
микросхемы.
Ceterum censeo Carthaginem delendam esse
(Карфаген должен быть разрушен)
Марк
Порций Катон
I
Справка
База
данных "Medline" (USA)
Ключевое слово "angiogenesis".
За период "No limit" обнаружено 4439 публикации, из которых за 1997 год - 654 публикации, в том числе из России - 2; за 1996 год - 722 публикации, в том числе из России - 3.
Ключевые слова "angiogenesis in vitro".
За период "No limit" обнаружено 143 публикации. Публикаций из России не обнаружено.
Для того, чтобы проверить насколько тщательно в базе данных "Medline" отслеживаются публикации российских ученых, был произведен поиск по ключевым словам: фамилии и инициалы крупных отечественных исследователей в области микроциркуляции. Было обнаружено, что практически все публикации этих авторов в центральной печати внесены в базу данных "Medline".
Если бы в России было много работ по проблеме ангиогенеза, то они наверняка были бы внесены в базу данных "Medline".
На основании анализа обнаруженных публикаций можно сделать вывод, что мимо России в очередной раз проходит крупнейшее научное направление исследований, связанное с решением ряда фундаментальных биологических и медицинских проблем, в том числе проблемы управление морфогенезом (гистогенезом и органогенезом), ключевым звеном которой является проблема управления ангиогенезом; проблемы управления ангиогенезом со стороны злокачественной опухоли и ее развитие в связи с этим; разработка новых методов лечения злокачественных новообразований путем управления ангиогенезом в последних.
Поражает
масштаб и технический уровень этих
исследований. Широко применяются
достижения молекулярной биологии,
клеточной биологии, иммунологии,
генетической инженерии, биохимии,
экспериментальной морфологии,
биохимии, электронной техники.
Создана целая индустрия
научно-технического обеспечения
этих исследований.
ОТКРЫТОЕ ПИСЬМО П. Р. М-ЧУ
15.07.96.
Добрый день П. Р.!
Обращаюсь к Вам по рекомендации А. В. Н., военпреда N-ского завода СМ, который просил меня при этом напомнить Вам о разговоре у Р. П. Н. (разговор был о "чипах"). Мне стало известно, что вы имеете определенное отношение к нашей отечественной "Кремниевой долине".
Я являюсь руководителем проекта Российского фонда фундаментальных исследований №96-04-50991 "Клеточная и тканевая инженерия эндотелия (Формирование в культуре эндотелия in vitro функционирующих саморазвивающихся капиллярных сетей)". Для успешной реализации проекта нам необходимо создать биомикроэлектронную систему (реактор), в которой можно будет запустить самопроизвольный процесс формирования функционирующих микрососудов и получить in vitro искусственное живое тканеподобное образование. "Сердцем" этого реактора должна быть интегральная микросхема, возможно, с объемной топологией, в которой должны быть как электронные коммуникации, так и гидродинамические коммуникации, в том числе активные, возможно, пьезоэлектрические элементы ("микронасосы"), система датчиков температуры, давления и т. д. Речь идет о создании принципиально новой интегральной микросхемы (процессора).
При успешной реализации этого проекта, может произойти биотехнологический прорыв и выход на совершенно фантастические рубежи, например, создание объектов, получивших в фантастической литературе название "киборгов", новых биоэлектронных тканевых и органоподобных протезов, биотехнотронных усилителей физических возможностей человека.
Мы ищем организации и творческие группы, умеющие разрабатывать интегральные микросхемы со сложной топологией, имеющие соответствующие технологии и мощности для производства опытных образцов таких микросхем. Мы хотели бы установить контакты с такими группами и проинформировать их более подробнее о нашем проекте и предложить им участие в этом проекте. Если Вы знаете такие группы, то не могли бы Вы, ознакомить их с нашей идеей. Я готов выехать в любое место, где к проекту будет проявлен интерес и сделать более подробный доклад и провести конкретные переговоры.
Если Вы нам поможете в установлении таких контактов, то мы будем Вам очень признательны.
Я примерно представляю ситуацию в "Кремниевой долине", но несмотря ни на что, я настроен оптимистично. То, что произошло с отечественной электроникой, весьма печально, но естественно и логично. Нельзя выиграть шахматную партию, повторяя ходы противника. Однако, я не думаю, что из поражения в одной партии следует поражение во всем матче. В конечном итоге, на марафонской дистанции побеждает не тот, кто первым вырвался вперед, а тот, кто первым пришел к финишу.
Как мне представляется, будущее в микроэлектронике за биомикроэлектронными гибридами, оптимально сочетающими в себе свойства живой и неживой природы. То, что чрезвычайно сложно создать, используя всю технологическую мощь современной человеческой цивилизации, природа "самопроизвольно" создала несколько миллиардов лет назад.
То, что я предлагаю, является, быть может, первой попыткой создания биомикроэлектронного гибрида.
На первых этапах этой НИР и ОКР нет необходимости в больших финансовых вливаниях, это все равно, что воду лить в песок. Нужны лишь хорошие, честные и бескорыстные головы, плюс немного разумного энтузиазма. Необходимо очертить круг лиц способных выполнить такую работу, провести с ними переговоры и приступить к разработке технических заданий по этому проекту. На эту часть работы средства, скажем так, есть. При разработке ТЗ задачи проектирования и технологии будут сформулированы предельно точно и ясно. Будут ясны ответы на вопросы: "Что, как, где, когда, сколько?". Я убежден, что после этого этапа некоторые государственные центры, имеющие высокую степень свободы выбора и толстые кошельки, не будут равнодушны к этой проблеме и ее решение можно будет перевести на уровень задачи государственной важности с соответствующей бюджетной поддержкой. Нам с Вами известны, по крайней мере несколько проектов, успешно решенных нашей страной в 40-е - 60-е годы подобным образом.
Ниже приводятся выдержки из указанного выше проекта:
1.5. Предполагается создать
экспериментальный реактор, для
культивирования эндотелия из вены
пупочного канатика человека,
обеспечивающего управляемое
включение в систему питательная
среда - эндотелиальная клетка
механического или
гидродинамического фактора в виде
организованных в микропространстве
тканевой культуры микропотоков питательной
среды. Предполагается что реактор
будет состоять из системы
жизнеобеспечения тканевой
культуры эндотелия,
генератора микропотоков
питательной среды, активной зоны
ангиогенеза, включающей устройство сопряжения
микропотоков с эндотелиальными
клетками, системы макро- и
микросенсоров, управляющего
компьютера с элементами
искусственного интеллекта, системы
визуализации процессов в
активной зоне ангиогенеза. Способность
эндотелиальных клеток в культуре
ткани in vitro образовывать
эндотелиальные сети с признаками начинающейся
канализации и установленное в
результате выполнения проекта
РФФИ 94-04-13544 "Структурный анализ микрососудистых
бифуркаций" значение
гемодинамического фактора для
формообразования микрососудистых
сетей in vivo позволяют
предположить, что организованное
включение гидродинамического
фактора в культуру эндотелия in vitro приведет
к формированию в последней
функционирующих саморазвивающихся
капиллярных сетей. Если
существование этого феномена
удастся доказать и его
воспроизвести, то фактически будут
созданы технологические
предпосылки для получения
искусственных биологических
тканей с заданными свойствами in vitro
4.3. Предполагается принципиально новый экспериментальный подход к изучению развития культуры эндотелия in vitro, характеризующийся включением в систему питательная среда - эндотелиальная клетка механического или гидродинамического фактора в виде организованных в микропространстве тканевой культуры микропотоков питательной среды. До сих пор при культивировании эндотелия in vitro решалась противоположная задача, а именно: исключить влияние на развитие эндотелия гемодинамического фактора, обязательное присутствие которого in vivo не позволяет изучить формообразующие потенции эндотелиальных клеток в чистом виде. Поставленная задача будет решаться путем создания принципиально нового устройства: реактора для культивирования эндотелия, который будет состоять из системы жизнеобеспечения тканевой культуры эндотелия, генератора микропотоков питательной среды, активной зоны ангиогенеза, включающей устройство сопряжения микропотоков с эндотелиальными клетками, систему макро- и микросенсоров, управляющего компьютера с элементами искусственного интеллекта, системы визуализации процессов в активной зоне ангиогенеза.
Проблема и задача, сформулированные в настоящем проекте, носят приоритетный характер и имеют абсолютную мировую новизну. Решение поставленных задач приведет к появлению технологических предпосылок для получения in vitro искусственных биологических тканей с заданными биологическими свойствами, что позволит принципиально по-новому решить проблему создания полноценных пищевых продуктов, проблему изготовления лекарственных препаратов, проблему создания полноценных биопротезов тканей и органов человека и животных и т. д.
В 1996 году предполагается провести теоретическое биофизико- и математико-морфологическое исследование клеточных (тканевых) систем человеческого организма, направленное на создание естественной классификации клеток человека, получить устойчивую культуру эндотелия из вены пупочного канатика человека и воспроизвести в ней все известные in vitro формообразовательные феномены эндотелия, выполнить их прикладную инженерную морфометрию, выполнить НИР и ОКР по указанным выше системам реактора, разработать технические задания, приступить к изготовлению действующего макета экспериментального реактора. При выполнении этапа 1996 года планируется максимально опираться на имеющиеся материально-технические ресурсы, широко использовать математическое моделирование для решения прикладных технических задач.
В 1997 году предполагается закончить изготовление и запустить макет экспериментального реактора в минимальной конфигурации, провести поиск оптимальных режимов формирования функционирующих и саморазвивающихся эндотелиальных капиллярных сетей in vitro, выполнить работы по совершенствованию технологии культивирования эндотелия, в основном закончить начатые в 1996 году теоретические исследования.
В 1998 году предполагается получить устойчивое формирование in vitro функционирующих и саморазвивающихся капиллярных сетей, начать на уровне НИР и ОКР разработку новых биотехнологий на основе этого феномена
4.4. Предполагается на основании теоретических биофизико- и математико-морфологических исследований клеточных (тканевых) систем, которые будут включать в себя компьютерный анализ накопленной информации о биологии, морфологии, физиологии конечных и промежуточных клеточных форм человеческого организма, как наиболее изученного из всех видов животных, построить или, по крайней мере, приблизиться к построению естественной классификации клеток человека, в некотором смысле аналогичной периодической системе элементов
Д. И. Менделеева, позволяющей проследить траекторию дифференцировки зиготы в каждую из известных конечных клеточных форм человека, получить устойчивую культуру эндотелия из вены пупочного канатика человека и воспроизвести в ней все известные in vitro формообразовательные феномены эндотелия, выполнить их прикладную инженерную морфометрию, выполнить НИР и ОКР с разработкой технических заданий по указанным выше системам реактора, изготовить их действующие макеты, приступить к изготовлению макета экспериментального реактора
4.5. В настоящее время известны автоматизированные технические системы для поддежания культуры тканей in vitro в течение длительного времени в проточной питательной среде, включающие датчики для отслеживания различных физико-химических параметров ее состояния и систем, поддерживающих постоянство ее состава.
Известно, что эндотелиальные клетки в культуре ткани in vitro самопроизвольно могут организовываться в сетевидные 3-мерные структуры, в результате соединения цитоплазматических отростков разных эндотелиальных клеток между собой, при этом в отдельных эндотелиальных клетках отмечается появление вакуолей, отмечается объединение вакуолей разных эндотелиоцитов. Известно также, что эндотелиальные сетевидные 3-мерные структуры в культуре ткани in vitro рано или поздно вновь распадаются на отдельные фрагменты с превращением эндотелиоцитов в клетки, напоминающие фибробласты. Эти феномены отражают формообразовательные потенции эндотелиальных клеток в чистом виде, без влияния гидродинамического (гемодинамического) фактора и фактически идентичны феноменам образования эндотелиальных капиллярных сетей in vivo в начальной фазе ангиогенеза, т. е. до момента канализации. Эндотелиальные клетки самодостаточны in vivo и in vitro для образования капиллярных сетей до момента их канализации.
Руководитель проекта считает, что для образования полноценных капиллярных сетей, т. е. для того чтобы прошла их полная канализация, как in vivo так и in vitro, необходим гидродинамический (гемодинамичекий) фактор. Настоящий проект направлен на доказательство этого положения. Ожидаемые результаты при успешной реализации настоящего проекта будут носить приоритетный характер и превосходить мировой уровень
4.6. Руководитель настоящего проекта в 1994-1995 гг. реализовал проект РФФИ 94-04-13544 "Структурный анализ микрососудистых бифуркаций". Результаты этой работы обобщены в монографии под названием "Структурный анализ микрососудистых бифуркаций. (Микрососудистый узел и гемодинамический фактор)" (Смоленск: АО "Амипресс", 1995. - 251 С.) [10 экземпляров монографии находятся в издательском отделе РФФИ]. Автором установлена формообразовательная роль гемодинамического фактора в ангиогенезе. Результаты этого исследования оригинальны, носят приоритетный характер и соответствуют мировому уровню новизны.
В состав основных исполнителей предлагаемого проекта введены специалисты, работающие в иммунологии, биохимии, механике, электоронике, математике и выполнившие в этих областях серьезные научные исследования, опубликованные в ведущих научных медико-биологических, технических журналах СССР, России и США, сделавшие 13 изобретений, реализовавшие ряд технических проектов для промышленности, обороны и космоса
4.7. Глотов В. А. Структурный анализ микрососудистых бифуркаций. (Микрососудистый узел и гемодинамический фактор). - Смоленск: АО "Амипресс", 1995
Теперь
некоторые соображения о контактах
с западом. Конечно, у них эту задачу
решать было бы проще. "Таньги"
много и комфорта тоже. Это, конечно,
в том случае, если они заметят и
оценят идею. Но с другой стороны,
деньги еще никого умнее не делали, а
комфорт расслабляет и
наркотизирует мозг. И за Державу
обидно. И вообще спортивный азарт.
Хочется их как следует "умыть".
С
уважением. В. Г.
II
ВАСКУЛОГЕНЕЗ И АНГИОГЕНЕЗ
Антология
(Как
это у нас начиналось)
"Глава, посвященная эндотелию, в настоящее время еще далеко не может считаться законченной, а представляет собой скорее поставку вопроса для дальнейшего исследования"
Н. Г. Хлопин
[Хлопин Н.
Г. Общебиологические и
экспериментальные основы
гистологии. - М.: АМН СССР, 1946.]
Термин "васкулогенез" употребляется для обозначения процессов возникновения сосудов из клеток предшественников в филогенезе и эмбриональном онтогенезе .
Термин "ангиогенез" употребляется для обозначения процессов развития сосудов из уже сформированных в эмбриональном, в постэмбриональном онтогенезе и при патологических процессах, в том числе при регенерации, росте опухолей, трансформации сосудов.
[Куприянов
В. В., Миронов В. А., Миронов А. А.,
Гурина О. Ю. Ангиогенез.
Образование, рост и развитие
кровеносных сосудов. - М.: НИО
"Квартет", 1993. - С. 7-8].
Фельдман Н. Г. М. Д. Лавадовский. - М.:Гос. изд-во мед. л-ры, 1956.
Первое исследование по проблеме ангиогенеза in vivo в России выполнено А.Е. Голубевым в работе "Материалы для анатомии, физиологии и истории развития волосяных сосудов" [Диссертация. - СПБ. - 1868]. Известна работа М. Д. Лавадовского "О развитии сосудов и нервов в хвосте головастика лягушки" [Журнал для нормальной и патологической гистологии, фармакологии и клинической медицины. - 1870. - Т. 1. - С.135-162].
Максимов А. А. Основы гистологии. Ч. I , II. - Л., 1925.
А. А. Максимов: "Уже у низших, беспозвоночных животных в теле часто имеется жидкость со взвешенными в ней различными клетками, кровь, либо находящаяся прямо в промежутках между органами и омывающая их, либо циркулирующая по вполне или частично замкнутой системе особых трубок, кровеносных сосудов. У позвоночных кровь разносится по телу всегда по замкнутой системе сосудов; необходимой, постоянной составной частью стенки последних являются располагающиеся в один слой плоские эндотелиальные клетки; в тончайших сосудах, капиллярах, кроме этого слоя эндотелия вообще ничего не имеется; в более широких сосудах, с более толстой стенкой, на эндотелий снаружи наслаивается еще разнообразные другие части - эластические волокна, гладкие мышцы и т. д.
При эмбриональном развитии всех позвоночных кровь впервые появляется уже очень рано, непосредственно вслед за формированием зародышевых листков и одновременно с образованием мезенхимы (Rckert и Mollier, 1906). Оказывается, что первые элементы крови - свободные, взвешенные в жидкости клетки - появляются вместе и одновременно с первыми зачатками сосудов и сердца - трубками, составленными вначале только одним тонким слоем эндотелия, и притом из одного общего источника. Этим общим источником, как для кровяных клеток, так и для эндотелия, сосудов источником развития и является именно мезенхима.
Сосудистая система появляется между зародышевыми листками не в одном, резко ограниченном участке, чтобы отсюда распространяться во все стороны, а всегда независимо, сразу или последовательно, во многих местах. При этом часть ее всегда и у всех позвоночных появляется уже с самого начала в виде сосудов, наполненных кровяными элементами, тогда как остальная ее область, между прочим как раз сердце и большие сосуды тела, закладываются в виде пустых тонкостенных эндотелиальных сосудистых трубок, в первое время совершенно лишенных клеточного содержимого и наполненных только жидкостью. Все это возникает отдельно, в разных местах зародышевого тела, вначале независимые друг от друга зачатки сосудистой системы, как наполненные кровяными клетками, так и пустые, затем быстро соединяются вместе; тогда получается сплошная, более или менее сложная, замкнутая сеть эндотелиальных соединительных трубок, один резко ограниченный участок которой функционирует в качестве сердца. Как только получилась такая объединенная система, кровяные тельца распределяются равномерно в жидкости, наполняющей все части ее, и начинается циркуляция.
У позвоночных с большими, богатыми желтком, меробластическими яйцами, а также у млекопитающих, где, как известно из эмбриологии, в тело зародыша превращается только небольшая часть яйца, остальная же внеэмбриональная масса последнего превращается в особый придаток зародыша, заключающий в себя желток, желточный мешок, первые, содержащие кровь, сосуды появляются в внеэмбриональной части яйца, в так называемом сосудистом поле (area vasculosa) и на поверхности желтка, где последний имеется (Rckert и Mollier, 1906; Minot, 1911; Максимов, 1907; Daranola, 1908; Keagan, 1917). Периферийный, продвигающийся между экто- и энтодермой край мезодермы не представляет собою здесь настоящего эпителиального слоя, а имеет целиком характер мезенхимы, состоя из неправильно разбросанных, отросчатых, подвижных, энергично размножающихся элементов. Когда начинается первое развитие сосудов и крови, эти элементы собираются в большей или меньшей величины компактные группы. Внутри последних клетки округляются и тесно сплачиваются, на периферии же их они остаются отросчатыми и такими же отросчатыми клетками, сильно вытягиваясь в длину, связывают соседние клеточные группы друг с другом, так часто получается сетеобразное расположение ткани. Описанные клеточные группы носят название кровяных островков. Клетки, лежащие на их периферии и в тяжах, связывающие отдельные островки, скоро уплощаются, соединяются своими краями, окружают круглые клетки и образуют в качестве эндотелия сеть тонкостенных сосудистых трубок . Круглые клетки, лежащие в глубине кровяных островков, наоборот, все становятся совершенно свободными, и между ними собирается жидкость - получается кровь, содержимое сосудистых трубок.
Тотчас вслед за образованием кровяных островков во внеэмбриональных частях зародыша, в его собственном теле, в щелеобразных промежутках между листками, вместе с обыкновенной мезенхимой, появляются на определенных местах отросчатые, подвижные сосудистые и сердечные клетки, образующие здесь пустые, лишенные клеточного содержимого сосуды с тонкой эндотелиальной стенкой и сердце. Все эти сосуды с самого начала всегда и со всех сторон окружаются мезенхимой. Во многих местах отдельные участки будущих сосудов появляются даже прямо в уже готовой мезенхиме, состоящей из сетеобразно соединенных друг с другом клеток, путем расширения некоторых межклеточных промежутков и уплощения окружающих последние клеток (H. von W. Schulte, 1914).
Но в
дальнейшем это очень скоро
изменяется. Образование новых
сосудов, постоянно требующееся при
быстром росте зародышевого тела,
впоследствии происходит уже не
путем новообразования из
мезенхимы, а только посредством
отростков, вырастающих из
предшествующих сосудистых
трубок".
***
Биографическая
справка
Максимов Александр Александрович
(15.02.1874-4.12.1928)
Родился в Санкт-Петербурге. С отличием окончил Петербургскую Военно-медицинскую академию (1896), до 1899 года работал там же на кафедре патологической анатомии, с 1900 по 1902 год стажировался во Фрейбургском университете. В 1903 году стал профессором Военно-медицинской академии, возглавил кафедру гистологии и эмбриологии. В 1922 году эмигрировал в США. С 1922 по 1928 годы профессор анатомии медицинского факультета Чикагского университета.
Основные научные труды. "Об ангиомах гортани" (1894), "Об изменениях паренхиматозных клеток печени при амилоидном перерождении" (1895), "К вопросу о патологической регенерации семенной железы" (1898), "О строении красных красных кровяных телец млекопитающих и о происхождении пластинок Bizzozero" (1898), "Экспериментальные исследования над воспалительными новообразованиями из соединительной ткани" (1902), "О воспалительных соединительнотканных новообразованиях у белых крыс..." (1903), "О клеточных формах рыхлой соединительной ткани" (1906), "Гистогенез соединительной ткани" (1909), "Лимфоцит как общая постоянная клетка различных элементов крови в эмбриональном развитии и в последующей жизни млекопитающих" (1909), "Исследования крови и соединительной ткани" ( 1909-1923), "О культивировании in vitro соединительной ткани" (1916), "Сообщение о клетках крови, соединительной ткани и эндотелия" (1924), "О способности развития лейкоцитов крови" (1925), "Основы гистологии" (1925), "О недифференцированных клетках крови и мезенхимных клетках зародыша во взрослом организме" (1926), "О развитии негранулярных лейкоцитов крови в полибласты (макрофаги) и фибробласты" (1927). А. А. Максимов автор унитарной теории кроветворения, ввел понятие "стволовая клетка", один из первых оценил метод тканевых культур и усовершенствовал этот метод (1916), дал классическую сводку по соединительным и кроветворным тканям (1927).
А. А.
Максимов во многом предопределил
направление развития мировой науки
в области клеточной биологии. Свои
взгляды, идеи, методы и подходы он
принес в США, где они хорошо
проросли и дали обильный урожай, в
том числе и Нобелевских премий. Его
труды стали мировой научной
классикой и до настоящего времени
остаются одними из наиболее часто
цитируемых среди работ
отечественных исследователей.
Литература:
Хлопин Н. Г. Проф. А. А. Максимов. -
Русский Архив анатомии, гистологии
и эмбриологии.- 1929.-Т. 8. - Вып. 1.- С. 3-6;
Мирский М. Б. А. А. Максимов (К
100-летию со дня рождения). - Проблемы
гематологии и переливания крови. -
1975. - Т. 20. - №6. - С. 53-57; Биологи.
Биографический справочник. Отв.
ред. Ф. Н. Серков. - Киев: Наукова
думка, 1984. - С. 394.
***
Щелкунов С. И. Интима мелких артерий и вен. // Архив биологических наук. - 1935. - Т. 37. - Вып. 3. - С. 609-637.
С. И. Щелкунов: "Данные эмбриогенеза сосудистой системы показывают общность происхождения элементов крови и сосудистой стенки. Кровь и сосуд - это два одновременно возникающих из одного и того же источника - мезенхимы, зачатка, в дальнейшем дифференцирующихся в различных направлениях. Однако связь между ними сохраняется на протяжении всей жизни организма. В процессе своей дифференцировки оба зачатка мезенхимы в различной степени теряют свойства исходной ткани. Часть их клеток в процессе развития получает новые морфологические качества и тем самым теряет способность к обратному метаморфозу.
Первичные сосуды, возникающие как в самом эмбрионе, так и в стенке желточного мешка, состоят вначале из одного слоя измененных мезенхимных клеток - первичного эндотелия, от которого в дальнейшем кнаружи развивается средняя и наружная оболочки стенки. Его принято считать за главную составную часть внутренней оболочки, хранящую в себе в течение всей жизни организма в значительной степени свойства мезенхимных образований. Эндотелий является источником соединительнотканных элементов и гладких мышечных волокон в интиме артерий. В культурах эндотелий мелких артерий, как показывают исследования Максимова, превращается в конечном итоге в обычных фибробластов. На природу эндотелия существует две совершенно различных точки зрения. Одна, как например Маршанд, Герцог, Тома и др. , считают, его за малодифференцированную клеточную генерацию. Маршанд в своих исследованиях приписывает эндотелию способность давать при делении адвентициальные клетки, гистиоциты и малые лимфоциты. Ф. Герцог показал фагоцитарную способность эндотелия капилляров, который ведет себя подобно макрофагам. Такого же мнения придерживаются Розенталь, Тома, Шульце и др. Наряду с этим существует другая точка зрения, которая была впервые высказана Максимовым, согласно которой эндотелий относится к числу высокодифференцированной клеточной генерации с весьма ограниченными возможностями к дальнейшему развитию и совершенно не способных давать ни адвентициальных клеток, ни лимфоцитов. Исключение составляют по Максимову лишь эндотелий капилляров печени, селезенки и костного мозга.
Однако
имеются и другие мнения, где за
эндотелием сохраняются некоторые
свойства мезенхимных клеток. Так А.
А. Заварзин в своем руководстве по
частной гистологии пишет
следующее: "Эндотелий
капилляров, а особенно
посткапиллярных вен не всегда
представляет собой какую-либо
законченную дифференцированную
клеточную форму"".
Щелкунов С. И. Прогрессивное и регрессивное развитие 1937. - капилляров. //Архив анатомии, гистологии и эмбриологии. - Т. XVII. - № 1. - С. 6-20.
С. И. Щелкунов. История вопроса (до 1937 года): "Развитие капилляров является одним из наиболее запутанных вопросов гистологии. Чтобы получить понятие о всей истории изучаемой темы автор начинает в хронологическом порядке с работ Келликера (Klliktr, 1846), Штриккера (Stricker, 1865, 1866), Голубева (Golubew, 1869), являющихся авторами одних из первых исследований, в которых наиболее полно представлен процесс развития капилляров.
Келликер исследовал развитие капилляров в оторочке хвоста головастика лягушки. Согласно его данным, развитие капилляров происходит за счет окружающих капилляр звездчатых клеток соединительной ткани. Последние своими отростками соединяются с конусообразными выростами капилляров, переходя постепенно в их стенку. Штриккер также наблюдал развитие капилляров в хвосте головастиков. Он считает, что развитие капилляров происходит путем отпрысков или тяжей, растущих от капилляров, которые вначале являются сплошными и только позднее внутри их образуется просвет для тока крови. Эти новообразованные капилляры представляют вначале простые протоплазматические трубки; клеточное строение развивается несколько позднее. Голубев на подобном же материале значительно дополнил описание Штриккера. Он отрицал наблюдения Келликера относительно участия соединительнотканных клеток в образовании новых капилляров. Вскоре сам Келликер вынужден был отказаться от ранее высказанного им взгляда. Голубеву не удалось ни разу наблюдать связь между сосудистыми отростками и соединительнотканными звездчатыми клетками. Развитие капилляров он описывает как разрастание в виде отпрысков клеток, расположенных на концах капиллярной сети, внутри которых постепенно образуется канал. Отпрыски, растущие навстречу друг другу, соединяются между собой. Присоединение к капиллярам из окружающей соединительной ткани новых клеточных элементов не происходит. Несколько позднее опубликованные исследования Арнольда (Arnolld, 1871, 1872) и Руже (Rouget, 1873) подтверждают данные Голубева и высказанный им принцип непрерывного разрастания капиллярной сети.
Далее необходимо остановиться на работах Ранвье (Ranvier, 1874), который дает принципиально новое объяснение процессу новообразования капилляров. В этом процессе Ранвье различает две стороны: разрастание ранее существующей капиллярной сети путем боковых отросчатых почек и развитие новой сети независимым от существующей и в дальнейшем связывающейся с общей сосудистой системой. Исследуя развитие сосудов в большом сальнике кролика, Ранвье отмечает, что в области млечных пятен развитие капилляров происходит при активном участии особых клеток, которые он назвал сосудообразующими или вазоформативными. Эти клетки являются постоянными элементами млечных пятен. По своей форме они отличаются от обычных соединительнотканных клеток. Они имеют отросчатую форму, часто принимающую вид узкого веретена с длинным ядром. Своими отростками клетки соединяются и образуют сети, которые Ранвье называет вазоформативными. В протоплазме этих клеток очень часто наблюдаются красные кровяные шарики, которые располагаются поодиночке или небольшими группами. На основании этих наблюдений Ранвье делает предположение о внутриклеточном образовании эритроцитов. К этому же положению пришел Шеффер (Scheffer, 1874) при изучении подкожной клетчатки крыс, где он встречал клетки, в протоплазме которых были заключены эритроциты. Однако эта точка зрения на эритропоэз не явилась для того времени новой. Она была высказана ранее Вебером (Weber), Герлахом (Gerlach,1849), Шаффнером (Schaffner, 1849) и др. в результате исследования зародышевой печени и селезенки, а в дальнейшем совершенно отвергнута Келликером.
Исследования Шпулера (Spuler, 1892) стоят в полном противоречии с данными Ранвье. В отношении новообразования капилляров он в значительной части подтверждает исследования Штриккера, Голубева и Арнольда, при этом совершенно отрицая существование вазоформативных клеток и сетей. Последнее, согласно с данными Шпулера, есть не что иное, как артефакты, получившиеся в результате разрыва капиллярной сети при искусственном растяжении ткани. На всех препаратах, при изготовлении которых не было искусственного растяжения, Шпулер наблюдал связь вазоформативных клеток и сетей с капиллярами. Это послужило основанием для его утверждений. Вместе с тем Шпулер отрицает возможность внутриклеточного образования эритроцитов, которая в то время допускалась Куборном (Kuborn), 1890, Николаидес (Nicolaides, 1891) и др.
В противоположность данным Шпулер, Франсуа (Francois, 1895) на основании изучения сосудов большого сальника кролика, подтверждает данные Ранвье о двух возможных источниках новообразования капилляров. Франсуа отмечает, что не подлежит никакому сомнению существование независимых от сосудистой сети вазоформативных клеток и сетей. Последние играют роль в возникновении новых капилляров, в дальнейшем присоединяющимся к общему сосудистому руслу. Кроме этого Фансуа указывает на гемопоэтическую роль вазоформативных элементов, в протоплазме которых образуются эритроциты. В отношении источника происхождения сосудообразующих клеток и сетей автор высказывает предположение. Что рассматриваемые элементы следует представлять как отделившиеся капсулы разрастания сосудов, при этом отделении их от общей сосудистой сети произошло по пути атрофии сосудов.
Восмер (Vosmer, 1898), изучив развитие сосудов сальника на эмбриональном материале кролика, выступил против образования капилляров in loco за счет вазоформативных элементов. Он считает, что вазоформативные сети с содержащимися в них эритроцитами есть не что иное, как обратно развивающиеся капилляры. Рено (Renaut, 1902) исследовал развитие сосудов в большом сальнике молодых морских свинок. Автор указывает, что в процессе развития сосудистой сети в некоторых ее звеньях происходит остановка в развитии, и в дальнейшем наступает атрофия этих участков. Жолли (Jolly, 1906) в результате изучения большого сальника у некоторых млекопитающих и, в частности , у котят и молодых крыс, приходит к следующему заключению: "Вазоформативная роль некоторых соединительнотканных клеток является очевидной: можно совершенно отчетливо установить, как периваскулярные, мезенхимные клетки удлиняются, располагаются в длинные ряды и образуют новые капилляры". В отношении независимых от сосудистой стенки вазоформативных сетей Жолли допускает две возможности. Или это искусственно отделенные сети, полученные при растяжении тканей. Или согласно данным Восмера и Рено, представляют собой фрагменты капиллярной сети, отделившиеся в результате атрофии участков, соединяющих их с сосудистой сетью. Заключенные в них эритроциты указывают на существующую ранее связь их с общей сосудистой сетью. Вместе с этим Жолли полностью отвергает внутриклеточное образование красных кровяных шариков. В дальнейшем данные Восмер, Рено и Жолли были также подтверждены исследованиями Мартынова (1907) и Парди (Pardi, 1909). Таким образом точка зрения, допускающая образование капилляров in loco за счет вазоформативных сетей, была совершенно отвергнута и восторжествовал снова взгляд Голубева и Арнольда, согласно которому капилляр происходит только от капилляра или, выражаясь словами Рабля: "Эндотелий происходит только от эндотелия".
На основании данных изучения эмбрионального развития первичных кровеносных сосудов из специального зачатка - ангиобласта [Гис (His), Майнот (Minot, 1908)], а также исследований, посвященных дальнейшему развитию сосудистой системы в эмбриональной и постэмбриональной жизни, принято до настоящего времени эндотелий выделять как особую клеточную генерацию с определенными морфологическими и функциональными признаками. На природу его существует два взгляда. Оба они указывают на специфичность эндотелия и расходятся лишь в оценке его степени дифференцировки. Одни рассматривают эндотелий состоящий из высокодифференцированных клеток, которые являются весьма ограниченными в своем дальнейшем развитии [Шпанг (Spang, 1936)], другие, наоборот, считают эндотелий за молодую генерацию, дающую в своем развитии различные клеточные формы, как например адвентициальные клетки, плазматоциты, моноциты, фибробласты и т.д. [Маршанд (Marchfnd), Герцог (Herzog), Тома (Thoma)]. Это превращение эндотелия считается необратимым. Подобно эндотелию, природа клеток наружного слоя капилляров является также неустановленной. Еще до сих пор некоторые исследователи [Вимтруп (Vimtrup), Крог (Krogh)] их считают за мышечные элементы, регулирующие капиллярное русло. Эта точка зрения еще до настоящего времени пользуется широким признанием среди работников клинической медицины. Другие исследователи (Маршанд, Герцог, Тома и др.) считают их за малодифференцированные клетки соединительной ткани. Беннингоф (Benninghoff, 1926) рассматривают их как часть фиброцитной сети, расположенной снаружи от капилляров. Этим самым он ставит знак равенства между адвентициальными клетками и обычными фиброцитами. Пфуль (Pfuhl, 1933) отрицает как мышечную природу адвентициальных клеток, так и их принадлежность к фиброцитам. Он относит их к особым клеткам, несущим важную роль в функции сосудов. Далее необходимо указать на весьма важные работы Кларков (Clark E. R. and Clark E. L.) и Сандисона (Sandison Y. C.), в которых при изучении капилляров применялся новый оригинальный метод исследования, дающий возможность наблюдать их при жизни. Полученные при этом данные расширяют существующее представление о структуре капилляров и выясняют функцию адвентициальных клеток. Несмотря на большое количество добытых фактов, морфология капилляров по настоящее время остается проблемой.
[Литература: 1. Arnold, J. - Virchow's Arch. Bd. 53 u. 54. 1871 u. 1872. 2. Benninghoff, A. - Zeitschrift f. Zellforschung u. Mikr. Anat. Bd. 4. S.125. 1926. 3. Clark, E. R. and Clark, E. L. - Americ. journ. of anat. Vol. 35. 1925; V. 49. 1932; V. 54, 55. 1934. 4. Dubovvik - Zeitschrift. Anat. Bd. 85. S. 178. 1928. 5. Essecke. - Klin. Wochenschrift. Bd. 2. 1923. 6. Francois P. - Archiv de Biologie. T.13. 1895. 7. Fuchs. - Anat. Hefte. Bd. 22. 1903. 8. Golubew. - Arch. f. mikr. Anat. Bd. 5. S. 49. 1869. 9. Gerlach. - Zeitschrift f. rat. Medizin. Bd. 7. 1849. 10. Herzog, F. - Klin. Wochenschrift. Bd. 3. S. 535. 1924. 11. Herzog, F. - Zeitschrift f. d. gts. exp. Med. Bd. 43. S. 79. 1924. 12. His. W. - Abhandl. d. mathem.- phis. Kl. d. Schs. Ges. d. Wiss. Bd. 26. S. 173-328. 1900. Leipzig. 13. Huzella, T. - Zeitschrift f. Zellforschung u. mikr. Anat. Bd. 2. S. 558. 1925. 14. Jolly. - Compt. Rend. Des Seances de la Soc. De Biol. 28. Juillet 1906. 15. Koelliker. - Annales des sciences naturelles. T. VI. 1846. 16. Kuborn. - Anat. Anz. Bd. 5. S. 277. 1890. 17. Loeschcke, Yans u. Elisabeth. Zeichrift f. microskop. anat. Forschung. Bd. 35. S. 535. 1934. 18. Marchand. F. - Mnch. Med. Wochenschrift. Bd. 70. S. 385. 1923. 19. Martinoff, V. - Internat. Monatsschrift f. Anat. u. Phisiol. Bd. 21. S. 281. 1907. 20. Minot. - Die Entstehung des Angioslastes und die Entwickl. d. Bluetes. Handbuch d. Entwickl. d. Menschen. Herausgegeb. v. Keisel u. Mall. Bd.2. 1911. 21. Nicolaides. - Arch. f. Anat. u. Physiol. 1891. 22. Pardi, F. - Arch. ital. di anat. e. di embriol. Vol. 8. Pag. 98. 1909. 23. Pfuhl. - Zeitschrift f. Zellforschung u. mikroskop. Anat. Bd. 20. 1933. 24. Rabl, C. - Morph. Jahrt. Bd. 12. 1887. 25. Ranvier L. - Archives de physiol normale et path. 1874. P. 429. 26. Renaut, M. J. - Compt. Rend. de l'assoc. des. Anat. Vol. P.63. 1901; Vol. 4. P. 230. 1902. 27. Rouget. - Arch. de physiologie. 1873. 28. Sandison, J. C. - Americ. Journ. of Anatomy. Vol. 41. P. 475. 1928. 29. Scelkunow, S. I. (Stchelkunoff, S. I.). - Zeitschrift f. d. ges. Anat. Bd. 103. 1934; 105. 1935; 106. 1936. 30. Scelkunow S. I. Archives d'Anatomie microscopique. T. 32. №1. 1936. 31. Schffer, E. - Proceedings of the Royal Society of London. Vol. 22. P. 243. 1874. 32. Schults, W. - Zeitschrift f. Anat. u. Entwickl. Bd. 76. S. 421. 1925. 33. Schwarz, G. - Virchow's Arch. Bd. 179. S. 209. 1905. 34. Spuler. - Arch. f. mikr. Anat. Bd. 40. S. 530. 1892. 35. Stricker, S. - Sitz. - Ber. d. kais. Akad. d. Wiss. Wien. 1865; 1866. 36. Vimtrup. - Zeitschr. f. Anat. u. Entwickl. Bd. 65. S. 150. 1922. 37. Vosmaer. - Verslag v. d. Gew. Vergad. d. Wid. Naturk. Afd. Deel 6. P. 245. 1898. 38. Zimmermann. - Zeitschr. f. Anat. u. Entwichl. Bd. 68. S. 29. 1923].
С. И. Щелкунов: "В развитии спайки следует различать три фазы. В течение первой фазы дегенерируют мезотелий и эластические образования. В их разрушении играют роль специальные лейкоциты и полибласты. Далее следует вторая фаза. В течение ее различно дифференцированные фибробласты, расположены по обеим сторонам эластических образований, претерпевают изменения. Связь их с окружающими промежутками и основным веществом нарушается. Они приобретают иную форму и красятся значительно интенсивнее. Вместе с фибробластами претерпевают изменения и попавшие сюда эксудативные полибласты и лимфоциты. В результате превращения всех означенных выше клеток образуется синцитий. Последний уже в ранних опытах является неоднородным. Внутренняя часть спайки слагается из длинных узких отросчатых клеток, с длинными овальными ядрами. Своими отростками они связаны между собой и принимают вид синцития. Обычно эти клетки красятся интенсивно железным гематоксилином. С обеих сторон этого слоя расположен синцитий иного характера, состоящий из более широких клеточных территорий, которые представляют собой неправильной отросчатой формы пластинки, переходящие одна в другую. Обе названные части синцития тесно связаны и образуют один общий пласт. Морфологическая неоднородность клеточных территорий синцития дает основание предполагать о их функциональном различии. Узкие, темноокрашивающиеся элементы, по-видимому, следует считать за недифференцированные мезенхимные элементы в противоположность более дифференцированным наружным слоем синцития. В третьей фазе происходит преобразование этого синцития. Образуется промежуточное основное вещество. Клеточные территории превращаются в клеточные элементы с ясно очерченным внешними контурами. В этой фазе возникают новые капилляры, которые, как показали опыты, появляются впервые на 4-6-й день после операции, в зависимости от скорости течения первых двух фаз развиваются спайки. Капилляры растут от краев лоскута к его середине. Вначале, как правило, они расположены лишь в недифференцированном отделе синцития, среди узких, интенсивно окрашенных элементов. Это обстоятельство указывает на взаимную связь между вновь развивающимися капиллярами и недифференцированными клеточными образованиями. В этом отношении изучение роста капиллярной сети дает более подробные данные. Полученные препараты показывают, что растущий конец капилляра всегда имеет связь с описанными выше длинными отросчатыми клетками. Эта связь не носит характера простого прилегания этих клеток к стенкам концов капилляров. Она имеет более близкую и при этом весьма своеобразную форму". Две рядом лежащие длинные отросчатые клетки, образующие недифференцированный отдел синцития, расположены на сохранившейся эластической сети. Их ядра с большим количеством хроматина имеют форму вытянутых стволов. В стенке растущего конца капилляра отмечается одна эндотелиальная клетка, имеющая овальное темноокрашенное ядро. Ближе к концу капилляра располагается вторая клетка, которая тесно связана с его стенкой. Эта клетка посылает протоплазматические отростки, которые постепенно теряются в окружающей ткани. Конусообразный конец капилляра связан с длинной отросчатой клеткой, которая является не чем иным, как составным элементом недифференцированного синцития. Растущий конец капилляра переходит в означенную клетку, часть которой превратилась в его стенку. "Другой свободный конец клетки сохраняется неизменным. От него в стороны отходят несколько боковых отростков. В протоплазме никаких изменений не отмечается. Просвет капилляра оканчивается большой вакуолью, отделенной от общего просвета тонкой продырявленной протоплазматической перегородкой. Эта вакуоль находится вблизи ядра клетки и содержит в себе эритроциты. Развитие просвета капилляра и вхождение означенных клеток в состав его стенки удается проследить на других участках препарата". Просвет растущего капилляра на сохранившейся эластической сети заполнен кровяными клетками. Конец капилляра связан с отросчатой темноокрашенной клеткой. Последняя с большим количеством боковых отростков одним своим концом незаметно переходит в стенку капилляра. Второй конец клетки лежит свободно. Просвет капилляра оканчивается несколькими различного размера вакуолями, расположенными в определенном порядке. Они лежат сбоку от ядра, оттесненного к периферии. Наиболее крупная вакуоль отделена от просвета тонкой протоплазматичесекой перегородкой, которая в средней своей части представляется разорванной. В просвете находятся кровяные элементы. Остальные вакуоли расположены в порядке уменьшения их размера, из которых самая меньшая находится ближе к свободному концу клетки, разделены между собой тонкими протоплазматическими перегородками. не содержат кровяные элементы. От стенки капилляра отходит в стороны значительное количество отростков, которые анастомозируют с отростками рядом лежащей одноименной клетки. "Вышеизложенные данные показывают, что при развитии капилляров большое значение имеют клетки недифференцированного отдела спайки. Этот отдел синцития является не только местом развития капилляров. Но его клетки являются элементами, из которых, по-видимому, и строится капиллярная стенка. Развитие капилляра, которое принято описывать со времени Штриккера, Голубева и Арнольда как размножение эндотелиальных клеток, расположенных в области конца капилляра на полученном автором материале ни разу не наблюдалось. Это, конечно, не дает права отрицать подобную возможность в развитии капиллярной сети. Возможно, она имеет место в других органах и тканях, в иных условиях их деятельности. Однако на материалах поставленных опытов этой формы развития видеть не удается. В нашем случае развитие капилляра протекало таким образом, что его стенка оказывалась преобразованной элементами недифференцированного синцития, в которых прокладывается кровяное русло. Последнее постепенно развивается, переходя от одной клеточной территории к другой. Этот переход завершается через протоплазматические связи, расположенные между отдельными элементами синцития".
Интересным является образование капиллярного просвета. Его развитию предшествует образование вакуолей в протоплазме клетки. "Обычно вблизи просвета растущего конца капилляра наблюдается одна или несколько вакуолей. Ближайшая к просвету вакуоль, достигнув определенного размера, соединяется с просветом капилляра. Протоплазматическая перегородка, отделяющая ее от кровяного русла исчезает. В результате этого, на внутренней поверхности стенки капилляра наблюдаются ее остатки, которые вдаются в просвет в виде небольших выступов. Таким образом русло капилляра удлинилось на величину одной, слившейся с ним, вакуоли. В дальнейшем происходит подобное явление со следующей, ближайшей к ней вакуолью. И так постепенно все вакуоли одна за другой присоединяются к кровяному руслу. Просвет удлиняется. В оставшейся части тела клетки идет образование новых вакуолей. В результате этого внутреннего преобразования протоплазмы клетки, вдоль ее тела образуется канал, заполненный кровью, который и представляет собой просвет новообразованного капилляра. Протоплазма клетки, растянутая тонким слоем становится капиллярной стенкой. Клетка из мезенхимальной превращается в эндотелиальную. Следовательно, просвет новообразованного капилляра не есть простая трубка, ограниченная плоскими, соединенными между собой. Эндотелиальными клетками. Он представляет собой внутриклеточное образование, возникшее в результате слияния большого количества вакуолей, расположенных в клетках мезенхимального синцития. Его можно рассматривать как одну весьма своеобразную, сложную "многоклеточную" вакуоль".
В 1874 г. Ранвье впервые наблюдал в сальнике молодых и новорожденных животных длинные, неправильной формы, отросчатые клетки, в протоплазме которых находились отдельные эритроциты. В 1895 г. подобные факты и на подобном материале наблюдал Франсуа. "Эти элементы названы Ранвье вазоформативными клетками (cellules vasoformatiives). Однако в двльнейшем подобные факты в работах Шпулера, Шварца. Мартынова, Парди и др. получили иную оценку. Описанные Ранвье и Франсуа вазоформативные клетки авторы рассматривали как элементы метаморфоза эндотелия в процессе обратного развития капилляров. Это предположение не находит для себя подтверждающих фактов. За то, что описанная выше картина преобразования, не является обратным процессом в развитии капилляров, а наоборот - является формой прямого развития, свидетельствуют два положения. Во-первых, внутрипротоплазматические преобразования клеточных территорий синцития наблюдаются в разных стадиях развития спайки, когда в ее средней части еще совершенно отсутствуют капилляры. В этих стадиях капиллярное русло растет со стороны краев лоскута в направлении к его середине. Во-вторых, описанные выше картины внутри протоплазматического преобразования наблюдаются лишь только в тех клеточных территориях синцития, которые непосредственно связаны с концами растущих капилляров. В других участках синцития их видеть не удается. Образование капилляров in loco с последующим присоединением их к общей сосудистой сети не наблюдается. Отсутствие митозов среди эндотелия концов капилляров, а также наличие вакуолей в протоплазме клеточных территорий синцития, свидетельствует о том, что их развитие происходит не путем отпрысков эндотелия, а в результате прокладывания капиллярного русла внутри протоплазмы клеточных территорий синцития. Возникшие таким образом новые капилляры в спайке брюшины следует назвать внутриклеточными (интрацеллюлярными) капиллярами, так как их просвет представляет внутриклеточный канал. Эта внутриклеточная форма сосудов является одной из начальных фаз в развитии капилляров. В дальнейшем, в результате деления клеток внутриклеточного капилляра стенка его приобретает клеточное. эндотелиальное строение. Просвет капилляра оказывается окруженным двумя, а иногда и более, эндотелиальными клетками. В следствие этого диаметр его значительно увеличивается. Капилляр из внутриклеточного превращается во внеклеточный (экстрацеллюлярный). Одновременно с этим прилежащие с наружи к эндотелию клеточные элементы синцития становятся обычными эндотелиальными клетками. В результате деления эндотелиальных клеток и одновременного превращения лежащих снаружи от эндотелия клеток синцития в мышечные элементы образуются прекапиллярные артериолы или посткапиллярные вены. На основании всего вышесказанного, в развитии сосудов спайки брюшины можно различать четыре фазы: в первой фазе происходит образование вакуолей в протоплазме клеточных территорий синцития, связанных с концами капилляров; во второй фазе в результате слияния вакуолей с капиллярным руслом возникают интрацеллюлярные капилляры; в течение третьей фазы наблюдается превращение интрацеллюлярных капилляров в экстрацеллюлярные; в четвертой фазе происходит преобразование некоторой части таких капилляров в прекапиллярные артериолы и посткапиллярные вены.
Не является ли развитие капилляров процессом обратимым, и если это имеет место, то как ведут себя различные клеточные элементы, составляющие стенку названных сосудов? В сосудах на различных стадиях их развития регрессивные изменения выражены неодинаково. Для каждого вида сосудов они имеют определенную морфологическую картину. В интрацеллюлярных капиллярах эти изменения наиболее резко выражены. Интрацеллюлярные капилляры разделяются на отдельные сегменты различной величины. Они занимают длину одной или нескольких клеток. В просвете их находятся кровяные элементы, которые в местах разрыва выходят в окружающую ткань. При регрессивном развитии капилляров капиллярная сеть распадается на отдельные фрагменты. Вследствие этого названные внутриклеточные пласты являются замкнутыми пространствами или в некоторых случаях, в результате разрыва клеток, открытыми в окружающую ткань. Вакуоли в протоплазме клеток, как правило отсутствуют. От полости отходят в различном направлении протоплазматические отростки, которые напоминают о ранее имевшейся форме клеток, вошедших в состав капилляра". Какова же дальнейшая судьба этих клеток и оставшейся в них части сосудистого просвета? Входят ли обратно в синцитий клеточные элементы интрацеллюлярных капилляров при регрессивном развитии? Сходство формы клеток капиллярного фрагмента с окружающими клетками свидетельствует о том, что в условиях регрессивного развития интрацеллюлярных капилляров их клетки входят в состав этой ткани. Таким образом о прогрессивном развитии интрацеллюлярных капилляров можно в некотором смысле говорить как о процессе обратимом. Дальнейшая судьба этих клеток невыяснена.
Регрессивное
развитие в экстрацеллюлярных
капиллярах протекает иначе. Общая
форма сети в большинстве случаев
сохраняется. Отмечаются лишь
отдельные разрывы, через которые
выходят кровяные клетки в
окружающую ткань. В большинстве
случаев капилляры находятся в
спавшемся состоянии. Просвет их
суживается, а в дальнейшем
совершенно исчезает.
Эндотелиальные клетки
превращаются в веретенообразно
вытянутые элементы. Вместе с
прилежащими к ним снаружи
адвентициальными клетками, которые
обычно вытянуты вдоль капилляра,
они образуют плотный тяж. Последний
может иметь различную толщину в
зависимости от количества клеток,
входящих в состав стенки капилляра.
Дальнейшие изменения этих клеток
проследить не удается, так как они
совершенно теряются в ткани спайки.
Что касается изменения стенки
прекапиллярных артерий, то оно во
многом напоминает изменения в
экстрацеллюлярных капилляров.
"Просвет артерии суживается и в
дальнейшем исчезает. Кровяные
клетки выходят в окружающую ткань.
Клеточные элементы стенки образуют
вместе толстый тяж. В некоторых
клетках эндотелия и
адвентициальных клетках
наблюдаются амитотические
перешнуровывания ядер и отдельные
митотические фигуры. По ходу этих
измененных артерий располагаются
отдельные полибласты, в
протоплазме которых скапливается
значительное количество
включений". Посткапиллярные вены
не образуют плотного тяжа, их
стенки становятся более рыхлыми,
слой эндотелия распадается на
отдельные отросчатые клетки,
которые вместе с наружными
клетками входят в состав
окружающей ткани.
Заварзин А. А. Курс гистологии и микроскопической анатомии. - Л.: Медгиз, 1938, 939. - С. 149, 289-293; Заварзин А. А. Очерки эволюционной гистологии крови и соединительной ткани. Избранные труды . В 4-х томах.- М.-Л., 1953.
А. А. Заварзин: "Эндотелиальные клетки во многом тождественны с молодыми, малодифференцированными фибробластами и даже с камбиальными клетками. Они сохраняют эпителиоидное расположение только в нормальных условиях; при различных же раздражениях они резко изменяют свой характер и приобретают вид молодых развивающихся фибробластов или даже камбиальных клеток, которые дальше могут дифференцироваться в том или ином направлении. В культурах эндотелий растет также отдельными тяжами, как и фибробласты.
Первые
сосуды зародыша возникают двояким
образом: в желточном мешке и в теле
зародыша. Образовавшиеся первые
сосуды, соединяясь вместе, дают
начало сети желточных капилляров,
которые вступают в сообщение с
сосудами, развивающимися в самом
теле зародыша. Эти последние
закладываются сначала прямо в
мезенхиме в виде щелей
неправильной формы, наполненных
жидкостью. Мезенхимные клетки,
образующие стенки этих щелей,
становятся плоскими и превращаются
в эндотелий, не утрачивая связи с
окружающей мезенхимой. Когда по
этим сосудам начинается
циркуляция, то очертания их
становятся более правильными, а
стенка, развивающаяся все время под
воздействием механических условий
кровяного тока, постепенно
приобретает все более сложное
строение, обнаруживающее ту
совершенную функциональную
приспособленность".
Хлопин Н. Г. Культура тканей. - Л., 1940.
Н. Г. Хлопин: "Культурой тканей (тканевой культурой, культурой in vitro или эксплантатом) называется частица, выделенная из целого многоклеточного организма и помещенная в среду подходящего состава, в которой возможны, при определенных температурных условиях, сохранение всех необходимых для жизни функций и увеличение массы живой протоплазмы, а вместе с тем рост и размножение клеток.
В богатых
капиллярами эксплантатах из
некоторых органов куриного
зародыша (У. Льюис, 1931; Тере, 1937),
сальника, подкожной ткани и
мозговых оболочек кроликов или
крыс (Максимов, 1916, 1925-1927;
Зильберберг, 1929; Герцог и Шоппер, 1931;
Скриба, 1935 и др.) описывается
прорастание сосудистых почек,
одновременно с ростом элементов
соединительной ткани. Льюис
наблюдал даже образование обширных
сетей капиллярных отпрысков.
Растущие капилляры имеют вид полых
внутри трубок разного калибра,
образованных тесно сомкнутыми
клетками эндотелия, с замкнутым и
большей частью заостренным концом.
Образуя боковые отпрыски,
эндотелиальные трубки могут в зоне
роста соединяться друг с другом. В
их просвете находится жидкость, в
которой бывают взвешены в разном
количестве кровяные клетки,
перетекающие пассивно из сосудов
центрального кусочка. Отпрыски
капилляров могут быть заметны в
зоне роста, по данным всех авторов,
лишь в течение нескольких дней
после эксплантации. В дальнейшем
они исчезают, частью погибая,
частью, по-видимому, распадаясь на
отдельные фибробластообразующие
элементы, неотличимые от
находящихся по соседству
соединительнотканных клеток. В
эксплантатах из соматической
мускулатуры новорожденных
кроликов автору приходилось видеть
появление в разных количествах
капиллярных отпрысков, часть
которых могла сохраниться до 10-15
дней в культурах, не подвергшихся
вырезанию".
Снесарев П. Е. Общая гистопатология мозговой травмы. - М., 1946.
П. Е. Снесарев: "Один из авторитетных исследователей сосудистых реакций Газа позволил себе следующее выражение: "Эндотелии имеют такую высоко развитую способность к регенерациям, как если бы они чувствовали себя ответственными за снабжение кровью погибающей ткани".
Первые
зачатки эндотелия появляются не в
мезенхиме тела, а вне его - в тех
ранних мезодермальных производных,
которые в виде эктобласта и
мезобласта участвуют в оформлении
полости целома. Гис указывал на
особый примордиальный источник их
образования - ангиобласт, который,
появившись во внеэмбриональной
части, затем врастает в тело
эмбриона".
Клосовский Б. Н. Развитие капилляров мозга. -М.:Медгиз, 1949.
Б. Н Клосовский: "....Вопрос, стоявший перед исследователями , которые занимались изучением роста капилляров. Был достаточно сложен. Предстояло выяснить из каких клеток образуются новые капилляры: из клеток уже существующих сосудов, или из клеток, находящихся вне капилляров, или из тех и других. В литературе до сих пор не различается достаточно четко врастание капилляров и сосудов в область, еще лишенную капилляров, от размножения капилляров на участке, уже имеющем сосуды. Большинство авторов, говоря о развитии капилляров, в действительности исследуют врастание, а не размножение их. Поэтому все пишут о "врастающих сосудистых отпрысках", благодаря чему создается представление, что все время в разные стороны от сосудистой сети растут слепо оканчивающиеся капилляры и эти слепые отпрыски занимают все новые и новые области. В действительности же, кроме этого периода врастания сосудов, необходимо различать период усиленного размножения капиллярной сети. Каким же образом происходит увеличение количества капилляров в этом случае? На этот вопрос в литературе автор ответа не нашел.
Ознакомление с работами, посвященными изучению роста капилляров в различных тканях, показывает, что по существу все авторы видели одни и те же картины. Но так как рост капилляров представляет собой динамический процесс и не развивается строго в одной плоскости, а, напротив, во всех плоскостях, то на тонких гистологических препаратах, хотя и взятых в разные сроки, нельзя видеть все отдельные части капилляров; вследствие этого отрывочные гистологические картины дают широкие возможности для различного их толкования.
.... Наиболее интересными являются данные Кларка (1939) [Clark E. R. a. E. L. Microscopic observathion on growth in liwig mammal. - Am. J. Anat. 64. 1939], наблюдавшего рост капилляров в оторочке хвоста у личинок амфибий через прозрачную камеру. В этой работе он описывает рост капилляров в виде протоплазматических тяжей, идущих от уже предшествующих капилляров. На переднем конце растущего капилляра имеется один или несколько толстых протоплазматических отростков, постепенно утончающихся к концу. Любопытны взгляды Кларка на клеточный состав растущего капилляра. Он категорически заявляет, что эндотелий происходит только от эндотелия, не приводя, однако, ни одной убедительной микрофотографии. Не совсем убедительны также утверждения Кларка (1939), что отпрыск капилляра растет между звездчатыми клетками соединительной ткани, но не вступает с ними в контакт, до тех пор, пока полностью не образуется капилляр, уже проходимый для тока крови (тогда звездчатые клетки как бы присоединяются к нему, образуя его адвинтицию). Кларк пишет: "...плотный конец растущего капилляра может иметь, а может и не иметь ядра".
Пути развития капилляров и отдельные его фазы. На капиллярах мозга эмбрионов и новорожденных располагающаяся с наружной стороны сосуда эндотелиальная клетка, вернее, ядро ее, становится более заметной, благодаря увеличению окружающей ядро протоплазмы. В это время дистальная часть протоплазмы вокруг ядра увеличивается и вытягивается в небольшой столбик, который начинает выпускать отростки. Общий вид такой эндотелиальной клетки становится похожим на распускающуюся почку (стадия "почки"). Что представляют собой отростки клетки в действительности, мы пока не знаем, но на импрегнационных препаратах они выявляются то в виде тонких эластичных веточек или ворсинок, то в виде маловетвистых более или менее оформленных протоплазменных отростков, отчасти похожих на ветвления гортеговских клеток. Иногда на отростках импрегнируются утолщения, напоминающие присоски. В дальнейшем та часть протоплазмы, которая соединяет ядро эндотелия с основным сосудом, все более увеличивается в массе, превращаясь в цилиндрические тяжи различной толщины. Ядро эндотелия с окружающей его протоплазмой. Которая на переднем конце представляется резко уплотненной, и с отходящими от этой уплотненной части протоплазмы отростками все больше отходит от основного сосуда. На этой стадии прорастания капилляр получает внешнее сходство с полипом, почему автор и называет эту стадию стадией "полипа". В этой стадии передний конец растущего капилляра булавовидно утолщен, интенсивно импрегнирован и отростки его значительно вытянуты. В отношении отростков необходимо заметить, что и на этой стадии они представляют собой сильно меняющиеся образования: чаще всего они имеют вид коротких ворсинок, волнообразно изогнутых, равномерных по всей длине, более или менее симметрично отходящих от уплотненного переднего конца протоплазмы, благодаря чему получается вид розетки. В следующей стадии своего развития вырост эндотелия продолжает увеличиваться в длину; он имеет цилиндрическую форму и характеризуется довольно плотной, хорошо импрегнирующейся протоплазмой. На переднем растущем конце, немного не доходя до ядра, на цилиндрическом тяже отмечается перетяжка, истончение веретенообразной формы (иногда этой перетяжки может и не быть). Ближе кпереди цилиндрический тяж обычно просветлен; в нем располагается ядро эндотелия; конец протоплазматического тяжа уплотнен, сильно импрегнирован и снабжен отростками, варьирующими в своем числе, форме и направлении; чаще всего они имеют вид щупальцев гидры, равномерно расходящиеся во все стороны, почему автор и называет эту фазу стадией "гидроида". Продолжая удлиняться, протоплазменный тяж приближается к соседнему сосуду; когда он подходит на близкое расстояние, часть отростков исчезает, а другие, наоборот, удлиняются и своими концами как бы оплетают сосуд или присасываются к его стенке. Наконец передний конец тяжа пристает к стенке сосуда и сливается с ним. Однако еще некоторое время отростки остаются видимыми на месте соединения конца капилляра со встреченным им сосудом. При изучении описанного выше хода развития капилляров в мозгу создается впечатление, что каждый капилляр является результатом превращения одной клетки эндотелия. При изучении препаратов капиллярной сети коры полушарий головного мозга взрослой особи, действительно на большинстве капилляров автор видел только одно ядро эндотелия. В этом отношении как будто капилляры мозга, состоящие из одной эндотелиальной клетки, отличаются от капилляров в других органах животного организма, где они состоят из большого числа преобразованных эндотелиальных клеток. По своей протяженности капилляры мозга значительно меньше, чем капилляры других тканей. В то же время среди мозговых капилляров встречаются и длинные, в которых можно обнаружить не одно, а несколько эндотелиальных ядер. Образование многоклеточных капилляров автор представляет себе на основании имеющихся у него препаратов как результат размножения и преобразования тех эндотелиальных клеток, которые сползают на растущий капилляр с основного сосуда. Таким образом, получается, что направляющая рост эндотелиальная клетка, находящаяся на переднем конце растущего капилляра, какой бы длины он ни был, является основной. Удлинение же капилляра достигается добавлением новых клеток эндотелия, отщепляющихся от основного сосуда. Можно полагать, что проникновение новой эндотелиальной клетки в протоплазматический тяж растущего капилляра дает импульс для дальнейшего его роста. Так, часто встречаются растущие капилляры, которые имеют ядро не только на своем переднем конце, но и посередине тяжа. В таких случаях возле второго ядра снова видны отросточки, отходящие в разные стороны, благодаря чему капилляр приобретает вид тяжа покрытого щетинками.
До настоящего времени остаются невыясненными потенциальные возможности эндотелиальной клетки сформировавшегося капилляра. Предстоит еще решить вопрос, сохраняет ли свои пролиферативные и регенеративные возможности эндотелий функционирующих, проходимых для тока крови капилляров. Если эндотелий сформировавшегося капилляра сохраняет способность к дальнейшему размножению, то при определенных изменениях среды, окружающей капилляр, может произойти образование нового капилляра из эндотелия уже функционирующего капилляра. Напротив, при утрате эндотелием сформировавшегося капилляра пролиферативных способностей образования новых капилляров можно ожидать только за счет эндотелиальных элементов больших сосудов, которые, как известно из патологии, сохраняют свои пролиферативные и регенеративные возможности.
Факторы, обусловливающие рост капилляров. При выяснении причин роста капилляров необходимо, во-первых, различать факторы, обусловливающие врастание капилляров в область, еще лишенную сосудов; во-вторых, выяснить, какие условия необходимы для размножения капилляров после их врастания в область, уже занятую сосудами. Действует ли при этом одни и те же факторы или природа их различна? Несмотря на то, что уже давно высказывались разнообразные предположения о возможных стимуляторах роста капилляров в разных органах и тканях, единого мнения по этому поводу получено не было, и, как справедливо замечает Кларк (1939), наши представления о факторах, обусловливающих рост капилляров, остается до сих пор в области предположений.
Роль сердечной деятельности и кровяного давления. Еще в 1898 г. Тома (Thoma) различал ранние примитивные васкулярные образования, появление которых обеспечивается врожденными факторами, и позднее образование новых капилляров, стимулом для которых служит начинающаяся сердечная деятельность и увеличение кровяного давления. Эта теория встретила в дальнейшем ряд возражений, прежде всего потому, что в ряде экспериментальных работ было показано, что образование новых капилляров может иметь место не только при торможении сердечной деятельности фармакологическими средствами [Стокард (Stockard, 1915)], но и при полном отсутствии циркуляции после удаления сердца у эмбриона [Петерсон (Peterson, 1909); Чапмен (Chapman, 1918); Кларк, 1918] и даже в культуре тканей [ Льюис (Lewis, 1931)]. Если считать возрастающее кровяное давление основным стимулятором новообразования капилляров, то наибольшее количество возникающих вновь капилляров нужно было бы искать на артериальных сосудах. И действительно, на своих препаратах из ствола мозга автор установил, что новые капиллярные выросты возникают преимущественно на артериальных стволах. В 1937 г. была высказана Вейраухом и Гарисом (Weyrauch u. Garis) точка зрения, близкая к мнению Thoma. Авторы полагали, что решающим стимулом для образования новых капилляров может являться увеличение объема крови и расширение капилляров. Однако это мнение вызывает совершенно справедливое возражение, ибо подобного рода гипотеза не может объяснить богатый рост капилляров с образованием многочисленных анастомозов и густой капиллярной сети не только в тканевых культурах, но и во всех случаях отсутствия циркуляции у эмбриона с удаленным сердцем. На основании приведенных противоречивых фактов можно сделать лишь следующий вывод: если высота кровяного давления и играет известную роль в новообразовании капилляров, то это все же не единственный определяющий его фактор.
Теории химической стимуляции роста. Ру (Roux, 1985), Маль (Mall, 1906), Вильямс (1937) высказали предположение, что повышенный метаболизм ткани является основным моментом, обусловливающим новообразование капилляров, поскольку возникновение сосудов наблюдается в эмбриональной ткани, находящейся в стадии кариокинетического деления. Это предположение, однако, недостаточно обосновано, так как усиленный кариокинез и новообразование капилляров, по-видимому, лишь сопутствующие друг другу явления. Леб (Loeb, 1893) и Ивенс (Evans, 1903) пытались объяснить рост новых капилляров, причем стимулом роста, по их мнению, должно быть какое-то химическое вещество. Выдвинув в 1918 г. гипотезу, ставившую рост и обратное развитие капилляров в прямую зависимость от обмена через эндотелиальную клетку, Кларк в ряде последующих работ (1919, 1932, 1935) делал попытки найти химические стимуляторы роста капилляров. Предпринятые с этой целью эксперименты, в которых было произведено инъецирование зерен крахмала, жировых капель, убитых культур бактерий, желатины и др. в-в, не дали положительных результатов. Рост капилляров от имеющихся уже кровеносных сосудов наблюдался лишь в поздних стадиях воспаления - после инъекции кротонового масла. На основании этого Кларк высказал предположение, что химический стимулятор роста капилляров нужно искать в воспалительных экссудатах или еще в эмбриональной ткани. Если принять, что химическую субстанцию, вызывающую рост капилляров, надо искать только в эмбриональной ткани и воспалительных экссудатах, то значительно ограничивается время и условия возникновения новых капилляров, так как приходится сделать заключение, что во взрослом организме, при обычных условиях, роста капилляров не происходит. В действительности же хорошо известно, что образование новых капилляров в различных тканях имеет место и у взрослых особей.
Ряд полученных автором фактов говорит в пользу того, что стимул для роста капилляров лежит во вне сосудистой стенки и химический агент, стимулирующий рост капилляров тем или иным путем, при тех или иных обстоятельствах, выделяется клеточными элементами мозговой ткани. Например, в полушариях головного мозга (отдел мозга, наиболее подходящего для анализа причин роста капилляров, так как области с повышенным метаболизмом здесь располагаются слоями) на первой стадии развития переднего мозгового пузыря капиллярные сосуды прорастают в виде радиальных, параллельно расположенных стволов, идущих через довольно широкий краевой покров, и развиваются в матриксе, где в это время происходит усиленный кариокинез материнских клеток. С началом миграции клеточных элементов на периферию мозгового пузыря и образованием коры отмечается усиленный рост капилляров в корковом слое, где происходит не только дифференцировка мигрировавших клеточных элементов в нервные и глиозные клетки, но имеет место и частичная их гибель. Можно предположить, что мигрирующие из матрикса на периферию мозгового пузыря клеточные элементы, не встретив здесь достаточного количества питательных веществ и кислорода, не только меняют свои обменные функции, но в некоторой части погибают. В таком случае как продукты измененного обмена, так и химические ингредиенты погибших клеток могут выступать в качестве химических стимуляторов. Обуславливающих повышенную деятельность сосудистого эндотелия. Явления асфиксии и моменты механической травмы при родах приводят если не к гибели части нервных клеток. то к повреждению значительного числа их и тем самым к изменению обмера поврежденных клеток. Продукты измененного обмена, а также продукты распада погибших клеток, задерживающихся в мозгу вследствие возникающего при этом отека и набухания мозговой ткани, могут являться агентами, стимулирующими рост капилляров. Для проверки этих предположений в лаборатории автора были предприняты эксперименты, ставившие своей целью выяснить действие асфиксии различной продолжительности на разные стадии роста капилляров (Киселева, 1948) [Киселева З. Н. Особенности роста капилляров при асфиксии. //Сборник рефератов работ Академии медицинских наук. 1948]. В ряде серий опытов было установлено, что длительное действие асфиксии ведет к прекращению роста капилляров, тогда как кратковременная асфиксия стимулирует их рост. Незначительная травма мозгового вещества новорожденных животных, ведущая к некрозу небольшой части нервной ткани, является стимулирующим моментом для роста капилляров не только в местах, прилежащих к некротическому участку, но и в противоположном полушарии (Космарская, 1948) [Космарская Е. Н. Объем, условия и последствия коллатерального кровообращения мозга. //Диссертация. 1948]. Полученные данные позволяют предполагать, что химические вещества, образующиеся в результате измененного обмена нервных клеток или их гибели (аноксия), являются стимуляторами роста капилляров.
...На основании изучения препаратов, полученных импрегнационной методикой автора, можно утверждать, что канализация растущего капилляра происходит после присоединения его к другому капилляру. Что светлое пятно на конце растущего капилляра представляет собой ядро эндотелия. А не просто вакуоли.
...Надо
решительно отвергнуть возможность
образования мозгового капилляра из
каких-либо других мезенхимных
элементов, как допускают некоторые
авторы для других тканей, так как
таковых в мозгу нет. Единственные
мезенхимные элементы, существующие
в мозгу, - гортеговские клетки -
являются частично продуктом
развития тех мезенхимальных
клеток, которые мигрируют в мозг
вместе с врастающими
капиллярами".
Клосовский Б. Н. Циркуляция крови в мозгу. - М., 1951.
Б. Н.
Клосовский (Метаморфозы): "...Не
существует единого мнения по
такому важному вопросу, каким
является вопрос о клеточном
составе растущего капилляра.
Разногласия настолько велики, что
даже в пределах одной и той же
гистологической школы уживаются
совершенно противоположные в этом
отношении взгляды (можно сравнить,
например, точки зрения А. А.
Заварзина и С. И. Щелкунова). ...За
счет чего же осуществляется рост
капилляров в замкнутой
сосудисто-капиллярной сети мозга?
Этот вопрос не новый, но по существу
возвращает нас к неоднократно
обсуждавшейся проблеме
дифференциации эндотелия. Можно
предположить, что появление нового
капилляра на стенке уже
сформированного, проходимого для
крови капилляра включенного в
замкнутую функционирующую
сосудисто-капиллярную сеть мозга,
происходит двумя путями. С одной
стороны, ядро растущего капилляра
может быть производным
эндотелиального ядра материнского
капилляра, претерпевшего деление.
Эта точка зрения должна
предусматривать
недифференцированное камбиальное
состояние капиллярного эндотелия,
способного давать многие генерации
при возникновении соответствующих
условий во внешней среде. [В своей
первой работе, посвященной
рассмотрению вопроса о
новообразовании мозговых
капилляров, автор был склонен
считать клетку строящегося
капилляра эндотелиальной (1949).
Однако более детальное
исследование этого вопроса
заставило автора отказаться от
такого взгляда]. С другой стороны
можно предположить, что
образование нового капилляра
осуществляется за счет особых
недифференцированных элементов,
входящих в состав сосудистой
стенки, тогда как эндотелий
исчерпывает свои возможности
размножения при построении лишь
одного капилляра. Следовательно,
надо полагать, что ядро эндотелия
заканчивает свою дифференцировку
после присоединения строящегося
капилляра к другому сосуду и
никакое обычное изменение во
внешней среде не может привести к
его делению. Клетка строящегося
капилляра при формировании
последнего способна давать начало
ряду новых капилляров. Ядро клетки
капилляра, организующегося в
функционирующей
сосудисто-капиллярной сети мозга,
способно к делению и не является
дифференцированным. Что же
представляет собой ядро этой
клетки по своей природе:
эндотелиальное ядро зрелого
сосуда, "сползающее с него после
деления с некоторым количеством
плазмы, или недифференцированную
клетку иного характера? Автор,
просматривая серии препаратов,
приготовленных из мозга животных
тех возрастов, в которых был
отмечен массовый рост капилляров,
не смог обнаружить никаких
признаков деления ядер
эндотелиальных клеток зрелых
капилляров. Независимо от размера
сосудов серого и белого вещества,
вплоть до капилляров, повсюду ядра
эндотелиальных клеток имеют
характерный вид, определенное
расположение хроматина и ничем не
обнаруживают возможности к
размножению. Вместе с тем на
наружной поверхности сосудов и
капилляров можно видеть ядра
других клеток весьма характерного
вида. Ядра этих клеток содержат
большое количество интенсивно
окрашенного хроматина,
расположенного в виде густо
переплетающихся толстых нитей или
в виде комков, лежащих плотно друг к
другу по длиной оси ядра. Клетки с
такого рода ядрами находятся не
только на радиальных сосудах коры,
где наблюдается особенно большое
количество и особенно тесное
расположение их на наружной
поверхности стенки, но и на
артериолах и капиллярах коры и
белого вещества. На последних число
ядер описанного вида значительно
меньше, расположены они могут быть
на капилляре в количестве одного
или двух. Уже одно характерное
расположение, вид и количество
хроматина говорят о
недифференцированном состоянии
ядер подобного рода клеток. На
недифференцированное состояние
этих ядер указывает также часто
наблюдающиеся в них различные фазы
деления. Характерным является факт
"сползания" клетки с описанным
ядром со стенки сосуда, на котором
она располагалась. В этих случаях
ядро отходит от сосуда на некоторое
расстояние и кажется отделенным от
него светлым промежутком
периваскулярного пространства.
Таким образом, клетки, которым
принадлежат эти ядра, являются
недифференцированными
образованиями, способными к
дальнейшему размножению,
обладающими возможностью
передвижения. Эти клетки, имеющие
источником своего происхождения
недифференцированные элементы
мягкой мозговой оболочки,
представляют собой не что иное, как
адвентициальные клетки, -
камбиальные элементы сосудистой
системы. В настоящее время,
основываясь на проделанных
наблюдениях, автор считает
строящийся капилляр мозга
результатом одного из превращений
адвентициальной клетки. Автор
считает, что весь
протоплазматический вырост
строящегося капилляра является
плазмой растущей адвентициальной
клетки, а не разрастанием
протоплазмы того капилляра, на
котором растет новый капилляр.
Стимулом для дифференциации этой
протоплазмы в новый капилляр
является неизвестное еще изменение
состояния тканевой среды,
окружающей капилляр, с
располагающейся на его стенке
клеткой адвентиции. Первым шагом к
дифференцированному состоянию
является выпускание протоплазмой
алвентициальной клетки
своеобразных отростков, что
происходит возможно, в силу каких
то изменений, наступающих к этому
времени в ядре этой клетки.
Адвентициальная клетка является
недифференцированным образованием
в течение всего периода роста
мозгового капилляра. Растущий
конец капилляра в свою очередь
может послужить для образования
двух и даже более новых капилляров.
Присоединение растущего капилляра
к другому капилляру означает
окончательную дифференцировку
адвентициальной клетки,
утрачивающей свои генеративные
возможности. Адвентициальная
клетка превращается в
эндотелиальную, а ядро ее - в ядро не
способного к дальнейшим
превраениям эндотелия. Описанный
способ организации вновь
образующегося капилляра в мозгу
является неизвестным до сего
времени".
Клосовский Б. Н. Мозг и его кровоснабжение. - М.:Знание, 1952. Серия II. - № 52.
Б. Н.
Клосовский: "...Развитие
мозговых капилляров идет
совершенно особым путем, отличным
от всего описанного до настоящего
времени в отношении роста
капилляров в других тканях.
Образование нового капилляра
начинается от уже существующего
капилляра, входящего в состав
замкнутой капиллярной сети мозга.
Адвентициальная клетка, лежащая на
материнском капилляре, начинает
увеличиваться в объеме и выпускает
в окружающую среду тонкие
протоплазматические отростки. Эту
первую стадию роста мозгового
капилляра назвали стадией
"почки"...".
Клосовский Б. Н. Развитие мозга ребенка. - М., 1954. - С. 27-39.
Б. Н.
Клосовский:"...Образование
новых капилляров происходит
следующим образом. На наружной
поверхности мозговых сосудов
обычно располагаются особые
незрелые клетки, получившие
название адвентициальных. Когда
начинается рост нового капилляра,
на уже сформированном и проходимом
для тока крови капилляре или сосуде
появляется небольшое возвышение.
При рассмотрении этого возвышения
оно оказывается той же самой
незрелой клеткой...".
Клосовский Б. Н., Космарская Е. Н. Деятельное и тормозное состояние мозга. - М., 1961.
Б. Н.
Клосовский и Е. Н. Космарская: "У животных,
имеющих переходный тип строения
сосудистой системы мозга или
организацию ее в виде непрерывной
сосудисто-капиллярной сети, в
определенной стадии развития на
смену процессу выгибания приходит
процесс роста мозговых капилляров
отпрысками. В ходе онто- и
филогенеза преобразование
сосудистой системы из конечных
артерио-венозных единиц в
непрерывную сосудисто-капиллярную
сеть происходит при участии
четырех процессов. Этими
процессами являются: процесс
слияния, выгибания, размножения
отпрысками и рост в длину. Среди
ряда неясных вопросов, касающихся
строения и развития сосудистой
системы мозга, нужно назвать вопрос
о способе врастания сосудов в
мозговое вещество. Известно, что
Стерци (1904) предполагал врастание
сосудов выгибанием. Напротив,
Вислони (1938, 1939) считал, что сосуды
врастают в мозг двумя рядом
расположенными
неканализированными плотными
тяжами. Образование
артерио-венозной единицы
происходит в мозговом веществе, где
дистальные концы параллельно
растущих сосудов почему-то
соединяются между собой.
Наблюдения Е. В. Капустиной над
развитием сосудистой системы
сетчатой оболочки глаза позволяют
автору считать способ выгибания
универсальным принципом врастания
сосудов, справедливым для всех
тканей. Врастание сосудов в
формирующийся мозг происходит на
тем более ранней стадии развития,
чем выше организованно данное
животное. В мозгу высших
млекопитающих врастание
артериоло-венозных капиллярных
петель способом выгибания по мере
развития сменяется способом
врастания отпрысками. Отпрыски
сосудов, входящих в состав
сосудисто-капиллярной сети внутри
мозга, направляются к поверхности
мозга и являются по всей
вероятности, венозными
капиллярами".
Куприянов В. В. Пути микроциркуляции. - Кишинев: Картя молдовеняскэ, 1969. - С. 142.
В. В. Куприянов: "Генез новых капилляров путем почкования, очевидно, есть основной путь перестройки капиллярного русла и васкуляризации новых территорий. Это довольно быстрый процесс. Zaken (Цит. по А. М. Чернуху, 1967), например, высчитал, что врастание новых сосудов в кожный трансплантат у белой мыши происходит со скоростью 5 микронов в час. Через 48 часов капилляры в регенерате кожи уха кролика (E. Clark и E. Clark, 1943) при повышенном давлении становятся артериолами, приобретая гладкомышечные клетки.
Всегда ли
капилляры представляют отростки
(отпрыски) сосудистых трубок?
Большинство исследователей
считает, что это так".
Щелкунов С. И. О закономерностях развития кровеносных сосудов в онтогенезе . //Всесоюзная научная конференция по возрастной морфологии. Ч. 1. - Самарканд, 1972. - С.170-171.
С. И. Щелкунов: "Наиболее ранним проявлением дифференцировки мезенхимы является образование крови и кровеносных сосудов. Гемопоэз и вазогенез обусловлены развитием и дифференцировкой обмена веществ, являющимся интегрирующим фактором развивающихся клеточных структур живого организма.
С возникновением на раннем этапе кровообращения, в дальнейшей дифференцировке тканей сосудистой стенки, кроме главного, названного выше фактора, играет важную роль гемодинамика, соответственно с которой постепенно развиваются свойственные артериям, капиллярам и венам тканевые структуры.
Капилляры
являются наиболее пластичными
отделами кровеносной системы,
способными к непрерывной
перестройке своей сетевидной
архитектоники, при этом происходит
частичное запустение старых и
образование новых капилляров".
Куприянов В. В., Миронов В. А., Миронов А. А., Гурина О. Ю. Ангиогенез. Образование, рост и развитие кровеносных сосудов. - М.: НИО "Квартет", 1993. - С. 25-30.
В. В. Куприянов, В. А. Миронов, А. А. Миронов, О. Ю. Гурина: "Сосудистая система в эмбрионе начинает развиваться тогда, когда процессы диффузии не могут обеспечить метаболические требования пролиферирующих клеток. На этой стадии экстраэмбриональное боковое скопление мезодермальных клеток (они носят название гемангиобластов и находятся в тесном контакте с энтодермой стенки желточного мешка) в висцеральной (спланхноплевральной) мезодерме начинает дифференцироваться с образованием изолированных кластеров клеток. Эти кластеры дают начало центрально расположенным кровяным островкам , которые формируют примитивные клетки крови (эритробласты). Клетки, расположенные по периферии, уплощаются и начинают детерминироваться в сторону образования эндотелиальных клеток . Эти клетки из прилежащих клеточных кластеров устанавливают контакты друг с другом с помощью отростков, а затем после слияния в монослой дают начало мелким желточным сосудам. Ангиобластические кластеры и тяжи, ведущие свое происхождение из мезодермы на боковых сторонах эмбриона, дают начало выстланным эндотелиальными клетками сосудам, лишенными кровяных островков [Risau W. In: The development of the vascular system. Eds. R. N. Feinberg et al. Issues Biomed. Basel. 1991. V. 14. P. 58-68].
Вторым шагом в развитии сосудистой системы является васкулогенез - развитие кровеносных сосудов из in situ дифференцирующихся эндотелиоцитов кровяных островков. Кровяные островки растут и сливаются, что приводит к образованию так называемого первичного капиллярного сплетения. Очень важным является вопрос, есть ли процесс васкулогенеза в самом теле зародыша. Последние работы с трансплантационными химерами и специфическими моноклональными антителами [Pardanaud L. Et al. Development. 1987. V. 100. P. 339-349; 1989. V. 105. P.437-485] убедительно продемонстрировали, что кровеносные сосуды действительно дифференцируются in situ в теле зародыша. Малочисленность внутриэмбриональных кровяных островков свидетельствуют о том, что увеличение числа эндотелиоцитов здесь достигается путем интенсивной пролиферации в большей степени, чем за счет дифференцировки in situ. Кроме того, дифференцировка эндотелиальных клеток in situ в теле зародыша происходит без параллельной конкурентной дифференцировки клеток крови [Risau W. Et al. In: Molecular Mechanisms in Cellular Growth and Differentiation. Academic Press. 1991. P. 207-222]. Во время формирования энтодермы сосудистая зона в эмбрионе подразделяется на две концентрические зоны: внутреннюю прозрачную (area pellucida), непосредственно окружающую эмбрион, и более наружно расположенную менее прозрачную (area opaca). Энтодерма этих зон отличается не только по своей структуре, но и по своему происхождению. Если непрозрачная зона происходит из зародышевой стенки, то прозрачная формируется путем инвагинации через первичную полоску. Граница между этими зонами прослеживается четко. В мезодерме сосудистой зоны, лежащей поверх энтодермы, можно заметить существенные различия в распределении сосудов между прозрачной и плотной зонами. В последней сосуды имеют множественные ветвления. В прозрачной зоне ветвлений почти не обнаруживается. Граница между двумя типами строения сосудистого русла соответствуют границе между областями энтодермы. С помощью моноклональных антител против ангиобластов и гемангиобластов перепелки QH1 показано, что механизмами формирования сосудов в плотной зоне являются как образование сосудистых выростов, так и удлинение сосудов без образования почек роста. Предполагается, что дифференцировка ангиобластов и васкулогенез связаны (или индуцируются) с энтодермой, тогда как ангиогенез является определяющим механизмом развития сосудистой системы тех тканей и органов, которые свободны от производных энтодермы. Однако проверка этого предположения при культивировании in vitro и на хориоалантоисной мембране не обнаружила ангиогенных свойств энтодермы сосудистой зоны [Flamme J. et al. In: IInd Int. symp. Grouth, differentiation and pathology of vascular endothelium. Schloss Ringberg. 1991.P.12].
Итак, спланхноплевральный листок индуцирует формирование 1) эндотелиальных клеток, 2) остеокластов красного костного мозга, 3) кровяных островков или групп гемопоэтических клеток в дорсальной брыжейке. Соматоплевра (эктодерма+мезодерма) или соматоплевральная мезодерма не обладает свойствами индуцировать образование гемопоэтических или эндотелиальных клеток. Способность индуцировать ангиогенез распределена широко по всей спланхноплевральной мезодерме, тогда как способность индуцировать гемопоэз ограниченна в эмбрионе близким соседством к дорсальной аорте от уровня приблизительно 10-го сомита и каудальнее [Dieterlen-Lievre F., Pardanaud L. In: IInd Int. symp. Grouth, differentiation and pathology of vascular endothelium. Schloss Ringberg. 1991.P.13].
Тип мезодермальных тканей, образуемых в изолированной эктодерме, зависит от концентрации так называемых вегетализирующих факторов . При высоких концентрациях формируются мышцы и хорда, а при низких - кровяные островки, кроветворные стволовые клетки и эндотелий. Индуцирующими потенциями обладают щелочной и кислый факторы роста фибробластов (ФРФ), трансформирующие факторы-бета 1 и 2 [Knochel W. et al. Blut. 1989.V. 59. P. 207-213].
Скопления и тяжи внутриэмбриональных клеток-предшественников эндотелиальных клеток появляются по бокам и близко от средней линии эмбриона и параллельно ей. Сначала между клетками образуется просвет, а затем формируется пара продольно расположенных сосудов - дорсальные аорты, которые затем соединяются с сердцем. Отток венозной крови в раннем эмбрионе обеспечивается передней и задней кардинальными венами, которые также развиваются in situ и анастомозмруют с сердцем с помощью капиллярных выростов. Вместе взятые, эти элементы формируют примитивную сосудистую систему раннего эмбриона. Пустые микрососуды в теле эмбриона анастомозируют с капиллярами в плотной зоне и вторично заполняются эритробластами из сосудов желточного мешка.
В желточном мешке эндотелиальные клетки и предшественники эритробластов имеют общее происхождение и называются гемангиобластами . Предшественники эндотелиальных клеток, которые не связаны с образованием гемопоэтических стволовых клеток, называются ангиобластами . В теле эмбриона скопления клеток, образующиеся в тесной связи с аортами, рассматриваются в качестве источника дифинитивных гемопоэтических стволовых клеток, однако до сих пор не ясно, имеют эти кластеры и эндотелиальные клетки общее или раздельное происхождение.
Процессы васкуло- и ангиогенеза идут параллельно как при образовании вне- и внутриэмбриональных кровеносных сосудов, так и в последующем при органогенезе. В отдельных частях зародыша: в боковой пластинке, головной мезодерме - формируются изолированные от имеющихся сосудов пузырьки, состоящие из эндотелиальных клеток. Во время роста сосудистые почки сливаются с этими пузырьками. Все это приводит к тому, что уже в течение первых двух - трех дней эмбрион трансформируется из бессосудистой структуры в организм, в котором каждая ткань обеспечена кровеносными сосудами и примитивными капиллярными сплетениями. На самых ранних стадиях развития зародыша все части бессосудистой внутриэмбриональной мезодермы могут стать источником образования эндотелиальных клеток [Wilms P. et al. Anat. Embryol. 1991. V. 183. P. 371-377]. Так, если пересадить эмбриональные стволовые клетки в брюшную полость мышей, то там образуются первичные капиллярные сплетения [Risau W. et al. Development. 1988. V. 125. P.471-478].
Факт образования кровяных островков доказан экспериментально in vitro. При выращивании в культуре ткани суспензии эмбриональных стволовых клеток, которые были получены прямо из мышиного бластоциста, происходило образование цистоидных эмбриональных телец, содержащих клетки крови. Этот процесс стимулировался сывороткой, полученной из пупочного канатика. Если пересадить эмбриональные стволовые клетки сингенным мышам интраперитонеально, то образуются более или менее дифференцированные кровеносные сосуды. А при пересадки цистоидных телец или агрегатов эмбриональных стволовых клеток на хориоаллантоисную мембрану, они вызывают стандартный ангиогенный эффект.
Доказано,
что ангиобласты, развивающиеся в
эмбрионе, полностью отделены от
желточного мешка [Reagan F. P. Anat. Rec. 1915. V. 9.
P.229-241].
Дифференцировка ангиобластов в
головной области эмбриона
ограничена околосетевой и боковой
мезодермой, так что цефалическая
область вначале свободна от
ангиобластов [Coffin D. J., Poole T. J. Development.
1988. V. 102. P.735-748]. Ангиобласты
найдены во всей
внутриэмбриональной мезодерме за
исключением хорды и прехордальной
пластинки в начале сомитогенеза.
Они являются высоко инвазивными и
движутся через эмбриональную
мезодерму во всех направлениях. Эти
подвижные клетки могу срастаться
друг с другом или внедряться в
эндотелиальные цисты или каналы и
тем самым способствовать удлинению
сосудов. Процесс миграции
стимулируется энтодермой. Места
формирования периферических
эндотелиальных тяжей определяются
окружающей мезенхимой [Noden D. M. In: The development of the
vascular system. Eds. R. N. Feinberg, G. K. Sherer, R. Auerbach.
Basel et al. 1991. P.1-24]. Синтетический
пептид GRGDS, напоминающий
фибронектин и содержащий
RGD-группировки, резко уменьшает
миграцию эндотелиальных клеток [Christ B. et al. Anat.
Embryol. 1990. V.181. P.333-339]. В процессе
эмбрионального развития миграция и
пролиферация предшественников
эндотелиальных клеток приводит к
экспрессии фибронектина, затем
эндотелиальные клетки начинают
секретировать ламинин и тем самым
инициируют дифференцировку
примордиальных сосудов [Risau W.,Lemon V. Dev. Biol.
1988. V. 125. P. 441-450].
Белок - строитель кровеносных сосудов. - За рубежом, 1985. -№50 (1327). - С. 20.
"Ньюсуик", Нью-Йорк: " После более чем десятилетних исследований группа ученых медицинского центра Гарвардского университета сообщила, что удалось выделить и получить в очищенном виде белок, "ответственный" за формирование кровеносных сосудов у человека. "Открытие способа образования органов оставалось предметом мечтаний ученых в течение многих лет, - говорит Берт Вэлли, руководитель исследовательской группы, - теперь это реальность". Открытие позволяет разобраться в механизме роста тканей.
Вэлли и его
коллеги назвали это вещество
ангиогенином. Они выделили его в
очень малом количестве из раковой
опухоли толстой кишки. Ангиогенин
обнаружен также в нормальной ткани
печени. Ученые показали, что он
стимулирует развитие капилляров
после инъекции в оплодотворенные
яйца кур и роговицу глаза кролика,
которые обычно не содержат
кровеносных сосудов. Исследования
показали, что ангиогенин
представляет собой белок,
состоящий из 123 аминокислот,
образующих единую цепь. Используя
методы генной инженерии, группа под
руководством Вэлли
реконструировала в лабораторных
условиях ген, который управляет
производством ангиогенина в
человеческом организме. Это
достижение позволит осуществить
производство белка с помощью
технологии рекомбинантных ДНК для
проведения дальнейших
исследований".
Белок , стимулирующий рост кровеносных сосудов. - Природа. - 1996. - № 4. - С. 107.
"Природа": "Биохимикам из Гарвардской медицинской школы (Бостон, США) под руководством Б. Волли (B. L. Vallee) удалось выделить в чистом виде и исследовать ангиогенин - белок, стимулирующий ангиогенез - процесс развития кровеносных сосудов,
Ангиогенин был выделен из питательной среды, на которой выращивалась культура карциномы человека, В очень малых количествах (3.5 пикомоля) белок вызывает рост кровеносных сосудов в роговице кроликов. Первичная структура ангиогенина весьма сходна со структурой фермента рибонуклеазы (гомология составила 35%), причем она включает аминокислотные остатки, существенные для каталитической активности рибонуклеазы. Авторы предполагают, что они определяют и ангиогенную активность нового белка.
В
лаборатории Волли был клонирован
человеческий ген, кодирующий
ангиогенин. Этот ген состоит из
непрерывной последовательности
оснований, т. е. не включает
интронов (некодирующих элементов
гена), что значительно облегчает
его выделение и генно-инженерное
использование для синтеза
ангиогенина. Это в свою очередь
будет способствовать выяснению
механизмов регуляции ангиогенеза [Biochemistry, 1985, v.24, №
20, p. 5480-5499. USA]".
Клесов А. А. Ангиогенин - белок, ускоряющий рост опухоли. - Наука и жизнь. - 1986. - № 11. - С. 64-65.
"Наука и жизнь":"В декабре 1974 года между Гарвардским университетом (США) и компанией Монсанто был заключен контракт, который газеты назвали крупнейшим в истории между частной фирмой и частным университетом. Согласно контракту, университет в течение 12 лет должен был вести поиск биологического агента, ответственного за васкуляризацию раковых опухолей, и в случае его обнаружения сделать детальную химическую и биологическую характеристику агента с целью разработки метода подавления его функции в организме. Компания в свою очередь, полностью финансирует эти исследования, обеспечивает их необходимыми материалами, реактивами, оборудованием, лабораторными помещениями и т.д. и в случае удачи оставляет за собой права на полученные результаты, включая патентные права, производство соответствующих медицинских препаратов и их коммерческую реализацию. Работы по поиску "ракового агента" возглавили два крупных американских ученых - Бэрт Вэлли, член Национальной академии наук США, тогда директор лаборатории биофизических исследований Гарвардской медицинской школы, и Джуда Фолкман, известный хирург, также из Гарварда. О результатах этого соглашения рассказывает профессор Анатолий Алексеевич Клесов (Институт биохимии имени А. Н. Баха АН СССР), который некоторое время работал в той лаборатории, где сделано открытие,
А. Клесов: "Соглашение Гарвард-Монсанто было секретным. Вскоре, в марте 1975 года, произошла "утечка" информации. Скандал удалось замять, и исследования снова засекретили. Прошло 10 лет, в конце 1985 года представители Гарварда организовали пресс-конференцию, и итоги работы были опубликованы как в научной, так и в массовой печати. Они оказались действительно сенсационными, Найден белок, который назвали ангиогенином ("порождающим сосуды" в переводе с греческого), ничтожные концентрации которого (пико- и фемтограммы, т.е. 10-12, 10-15 г) вызывают быстрое образование в тканях густой сети тончайших кровеносных сосудов - капилляров. Поскольку ангиогенин выбрасывается в ткани раковой опухолью, то кровеносные сосуды будут направляться именно к опухоли, обеспечивая ее питание кислородом и другими необходимыми для роста веществами. Увеличение опухоли усиливает выброс ангиогенина, что, в свою очередь, ускоряет рост сосудов и подачу крови к опухоли, и так далее - до возникновения лавинообразного и катастрофического для организма процесса метастазирования.
Группа Берта Вэлли, который за это время стал директором Центра биохимических и биофизических исследований при Гарвардской медицинской школе, получила ангиогенин в высокоочищенном виде, определила его полную аминокислотную последовательность, выделила ген, кодирующий синтез ангиогена в человеческом организме. И установила полную нуклеотидную последовательность гена.
Неожиданно
оказалось, что ангиогенин,
состоящий из 123 аминокислотных
остатков, по структуре весьма похож
на человеческий фермент
рибонуклеазу. Однако сходство это
чисто внешнее, так сказать,
"портретное", а
функционального "родства" нет:
роль рибонуклеазы состоит в
расщеплении РНК, ангиогенин же, как
показали авторы открытия, этого не
делает, а рибонуклеаза в свою
очередь не обладает ангиогенной
активностью. В частности, она не
вызывает роста кровеносных сосудов
в роговице глаза кролика (с помощью
этого теста в группе Велли
проверяют ангиогенную активность
различных препаратов). Дело в том,
что в нормальных условиях роговица
глаза кролика не содержит
кровеносных сосудов, кроме лишь
тех, что находятся в кайме глаза.
При внесении ничтожно малых
количеств ангиогенина в роговицу
она довольно быстро покрывается
сетью мельчайших капилляров.
Густота и скорость их
возникновения может служить
количественной мерой активности
введенного препарата. Активность
же ангиогенина просто
поразительна. Как уже отмечалось,
он вызывает образование
кровеносных сосудов при крайне
малых концентрациях: одна часть на
квадриллион (1:1015). По замечанию
одного из авторов открытия. Это
эквивалентно доле одной десятой
цента в общем бюджете США.
Следующий этап исследований Вэлли
и его сотрудников заключается в
наработке достаточного количества
ангиогенина для проведения
клинических испытаний. Пока в
распоряжении Вэлли - около 0.1 мг
ангиогенина человека, но он
надеется с помощью генетической
инженерии клонировать ген белка в
подходящих бактериях или дрожжах и
получить соответствующий штамм -
сверхпродуцент ангиогенина".
В мире науки 1989. - № 10. - С. 13.
"В мире науки": "Дж. Фолькмэн из Медицинской школы Гарвардского университета, известный своими работами по ангиогенезу, отмечает, что многие раковые опухоли достигают заметных размеров только после того, как развивается система их кровоснабжения. Недавно в журнале "Nature" опубликовано описание экспериментов Фолькмана, из которых следует вывод, что появление способности стимулировать ангиогенез является критической стадией в развитии злокачественных новообразований.
Фолькмэн вместе с Д. Ханааном из Колд-Сприт-Хпрборской лаборатории и другими исследователями изучал развитие рака, используя новый модельный объект - животных, у которых путем генетической инженерии создана предрасположенность к определенному виду рака. В данном случае опыты проводились на мышах, у которых в клетках поджелудочной железы, производящих инсулин, экспрессируя ген, обусловливающий развития рака, вследствие чего эти клетки часто претерпевают злокачественную трансформацию.
В поджелудочной железе у таких мышей помимо опухолей имеются островки клеток в "предраковом" состоянии, характеризующиеся гиперплазией. Клетки в этих островках делятся быстрее нормальных, но их рост еще не полностью вышел из-под контроля. Исследователям удалось выделить и культивировать гиперпластические островки, а также опухоли и выращивать их в геле вместе с эндотелиальными клетками. При этом опухоли неизбежно побуждали близлежащие эндотелиальные клетки формировать структуры, подобные капиллярам (Вероятно опухоль выделяет вещества, способствующие росту кровеносных сосудов). Оказалось, что некоторые гиперпластические островки тоже обладают свойством вызывать ангиогенез. Число гиперпластических островков, способных индуцировать образование капилляроподобных структур, через 7 недель культивирования резко возрастало. Существенно, что параллельно возрастало и число опухолей, способных к стимуляции ангиогенеза.
Из этого
следует, что стимуляция
кровоснабжения - возможно, в
результате генетического
изменения - не просто важна для
роста опухоли, но является одним из
ключевых событий злокачественного
перерождения ткани. Если Фолькмэн
прав, подавление ангиогенеза может
стать эффективным способом борьбы
с развитием опухолей на ранних
стадиях, а также с другими
патологическими явлениями,
включающими образование
кровеносных сосудов, в том числе с
неоваскулярной глаукомой и
ревматоидным артритом. Фолькмэн
недавно сообщил в журнале
"Science" о том, что некоторые
сложные синтетические
углеводороды могут в 100-1000 раз
усиливать активность стероидов,
подавляющую формирование
кровеносных сосудов. Он отмечает,
что глазные капли, в состав которых
входят вещества, подавляющие
ангиогенез, препятствуют росту
кровеносных сосудов в роговице,
который бывает причиной слепоты
".
Куприянов В. В., Миронов В. А., Миронов А. А., Гурина О. Ю. Ангиогенез. Образование, рост и развитие кровеносных сосудов. - М.: НИО "Квартет", 1993. - С.130-132.
В. В. Куприянов, В. А. Миронов, А. А. Миронов, О. Ю. Гурина: "Развитие техники культивирования клеток и тканей вне организма позволило поставить вопрос о моделировании ангиогенеза in vitro. Вначале удалось вырастить в виде тканевой культуры эндотелиальные и гладкомышечные клетки крупных сосудов. В дальнейшем были выделены эндотелиальные клетки из микрососудов, перициты и другие клеточные компоненты сосудистого русла. Разработаны методы выделения эндотелиоцитов из строго определенных сегментов сосудистого русла с помощью возвратно-поступательной перфузии сосудов взвесью стеклянных шариков, диаметр которых соответствует диаметру изучаемого уровня гемоциркуляции.
Культивирование эндотелиальных клеток вне организма дает исследователю уникальную возможность изучать их поведение в наиболее "чистом" виде, без влияния трудно учитываемых в экспериментах in situ гемодинамических, нейрогуморальных факторов, исключив (или моделируя) взаимодействие между эндотелиоцитами и клетками других типов. Культура эндотелиальных клеток представляет собой уникальную модельную систему для анализа механизмов ангиогенеза.
Образование капиллярных выростов в тканевых культурах человеческого красного мозга впервые описано в 1953 году Woodard W. C. и Pomerat C. M. [Anat. Rec. 1953, 117, p. 663-683]. Эти выросты идентифицировались на третий день после посева, но уже на 10-12-е сутки они подвергались дистрофическим изменениям. Вначале эндотелиальные клетки вытягивали небольшие отростки по направлению к периферии, затем формировались сплошные клеточные тяжи, которые на поздних стадиях обнаруживали просвет, не содержащий клеток крови. Сплетения капилляров начинали формироваться между 1-м и 5-м днями после начала культивирования. Этот процесс продолжался в течение почти двух недель.
При сокультивировании фрагментов микрососудов с миофибробластами удалось получить образование тяжей эндотелиальных клеток и формирование почек сосудистого роста [Sato N. at al. Connective Tissue. 1987, v. 20, p.101-104]. Колечки аорты, заключенные в коллагеновый и особенно в фибриновый гель и культивируемые в бессывороточной среде, вызывают образование ветвящихся микрососудов. Этот ответ блокируется гидрокортизоном и стимулируется средой, кондиционированной клетками карциномы. Культивирование вывернутых наружу фрагментов аорты под слоем внеклеточного матрикса без добавления сыворотки в среду культивирования приводит к образованию на месте эндотелиального пласта капилляроподобных трубчатых структур [Nicosia R. F, Ottinetti A. Lab. Invest. 1990, v. 63, p. 115-122].
Впервые in vitro феномен ангиогенеза на клеточной системе удалось воспроизвести в 1980 году [Folkman J., Haudenschild C. Nature, 1980, v. 288, p. 275-288]: эндотелиоциты микрососудов кожи человека формировали трубчатые конструкции в среде кондиционированной опухолевыми клетками и клонированными эндотелиальными клетками аорты. Через 20-40 дней после посева в 50% образцов на границе между центром и периферией колонии образовывались капилляроподобные трубки. Клетки имели все характеристики капиллярного эндотелия и росли в отсутствии других клеток (фибробласты, тучные клетки), которые обычно необходимы для ангиогенеза in situ. Вначале внутри эндотелиоцита появлялась цилиндрическая вакуоль, затем вакуоли соседних клеток сливались, образуя трубки, связывающие эти эндотелиоциты. После того, как соединялись 4-5 эндотелиоцитов, появлялась густая сеть капилляроподобных трубочек. Трубки были заполнены частично фибриллярным, частично мембранозным, частично аморфным материалом, который затем нередко рассасывался. Просвечивающая электронная микроскопия показала, что трубка окружена цитоплазмой эндотелиоцита.
Данные наблюдения свидетельствуют о том, что информация, необходимая для образования капиллярных трубок и всей капиллярной сети, экспрессируется эндотелиоцитами. Другие клетки для этого не нужны. При обычных условиях культивирования клетки из тех же источников лишь образовывали монослой [Folkman J. Nature. 1980, v. 288, p.551-556].
Было обнаружено, что если эндотелиальные клетки из вены пупочного канатика человека выращивать на субстрате, покрытым человеческим фибронектином, с добавлением в среду культивирования гипоталямического фактора роста, а в следующих пассажах лишить их этих специфических добавок, то через 4-6 недель эндотелиоциты образуют трубки, одним концом прикрепленные к субстрату. По времени появления вторичного типа роста обычно совпадает с усилением выработки эндотелиальными клетками простациклина [Maciag T. et al. J. Cell. Biol. 1983, v.97, p.153-165].
Формировать трубчатые структуры оказались способными эндотелиальные клетки, выделенные из микрососудов надпочечников. При культивировании эндотелиоцитов микрососудов на коллагене IV-V типа эндотелиальные клетки образовывали капилляроподобные трубчатые структуры, имеющие просвет и хорошо организованные межклеточные контакты. При выращивании тех же клеток в трехмерном коллагеновом геле также появлялись трубчатые структуры. Для этих целей субконфлюэнтный слой эндотелия микрососудов, выращиваемый на коллагеновом геле, покрывался еще одним слоем коллагенового геля. После чего происходила ретракция монослоя, приводившая к возникновению сети ветвящихся и анастомозирующих тяжей клеток с формирующимся просветом. В просвете обнаруживался клеточный детрит - субстанция, похожая по структуре на материал базальной мембраны [Mardi J.A., Pratt B. M. J. Histochem. Cytochem. 1986, v. 34, p. 85-91].
In vitro
удалось сформировать трехмерную,
состоящую из сосудоподобных трубок
сеть, образованную эндотелиоцитами
микрососудов мышечной и жировой
ткани. Для этих целей кусочек
мышечной или жировой ткани
заключался в фибриновый или
коллагеновый гель. Через день
появлялись короткие
эндотелиальные тяжи, которые
вырастали из ткани и быстро
удлинялись. На 5-6 день в геле
образовывалась трехмерная сеть
анастомозирующих клеточных тяжей,
в которых можно было рассмотреть
просвет. Эндотелиальные трубки
непрерывны. На базальной
поверхности выявляется материал,
сходный по строению с веществом
базальной мембраны [Montesano R. et al. J. Cell. Biol.
Int. Rep. 1985, v. 9, p. 869-875; Montesano R. et al. J. Cell.
Biol. 1987, v. 97, p. 1648-1652]".
Справка
База
данных "Medline" (USA)
Ключевое слово "Folkman J.".
За период
"No limit" обнаружено 247
публикаций.
Куприянов
В. В., Миронов В. А., Миронов А. А.,
Гурина О. Ю. Ангиогенез.
Образование, рост и развитие
кровеносных сосудов. - М.: НИО
"Квартет", 1993. - С.153: "Мы
рассматриваем наш скромный труд
всего лишь как короткий интервал на
длинном марше в сторону крупнейших
открытий, авторы которых (а одним из
которых несомненно должен быть J.
Folkman) скорее всего получат
Нобелевскую премию".
III
РЕАКТОР
Цель
исследования
Изучение влияния механического
или гидродинамического фактора в
виде организованных в
микропространстве тканевой
культуры микропотоков питательной
среды на формирование полноценных
или функционирующих и
саморазвивающихся эндотелиальных
капиллярных сетей in vitro.
Концепция
реактора для запуска в культуре
эндотелия in vitro процесса
формирования функционирующих
саморазвивающихся капиллярных
сетей
Основной
элемент такого реактора должен
состоять из гибридной интегральной
электронной биогидромеханической
микросхемы нового типа (рис.1),
включающей замкнутую систему
разветвленных микроканалов (1а, 1в),
имитирующих реальную
микрососудистую сеть, для
циркуляции жидкой фазы питательной
среды, топология

Рис.
1
которых
соответствует топологии реальных
микрососудистых сетей, выполняющих
in vivo аналогичные функции; и
сопряженные с ними электронные
функциональные блоки управления
(5а, 5б). Стенки микроканалов должны
представлять из себя микронасосы -
активные электромеханические
элементы (3а, 3б, 3в), в основе работы
которых лежит пьезоэлектрический,
или магнитострикционный, или
электрострикционный эффекты;
которые способны изменять
внутренний просвет каналов под
действием внешнего
электромагнитного поля, либо
генерировать внешнее
электрическое или магнитное поле
при изменении гидродинамических
параметров циркулирующей в них
жидкости, в последнем случае
микронасос будет выполнять функцию
тензодатчика (2а, 2б, 2в). Микронасосы
должны включать клапанные
устройства, имитирующие
аналогичные устройства в реальных
микрососудах, позволяющие
запускать и поддерживать на
заданном уровне в течение
необходимого времени процесс
микроциркуляции жидкой фазы
питательной среды в системе
микроканалов микросхемы. С
системой микроканалов должны быть
сопряжены специальный резервуар
для механической оксигенации
жидкой фазы питательной среды (4а) с
системой микроканалов, подающих
кислород, фильтрационная и
адсорбционная микроколонка для
очистки и детоксикации жидкой фазы
питательной среды (4б). Система
микроканалов. должна включать
специальную зону сопряжения с
эндотелиальными клетками (7),
состоящую из капиллярных
микротрубочек с пористой стенкой,
выведен-
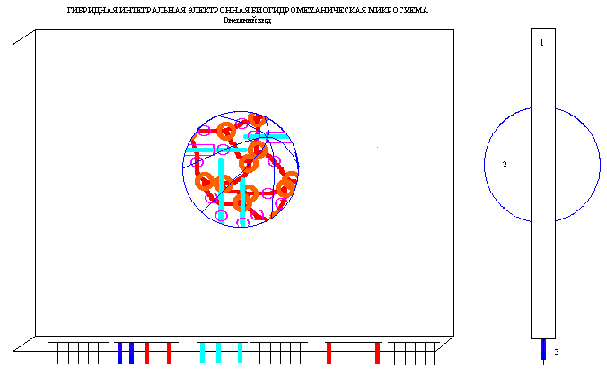
Рис.
2
ных в
специальный герметичный
микроконтейнер (рис.2),
монтирующийся в корпусе микросхемы
(1), заполненный желеобразной фазой
питательной среды. При помощи (рис.
1) термодатчиков (6а), термоэлементов
(6б), тензодатчиков (2а, 2б, 2в),
сопряженных с электронными
функциональными блоками,
производится контроль и управление
микроциркуляцией жидкой фазы
питательной среды в микросхеме и
поддерживаются необходимые
гемодинамические и температурные
условия для культивирования
эндотелиальных клеток. Все
основные функциональные элементы
микросхемы должны, по возможности,
создаваться на однородных
монокристаллах с использованием
современных достижений
микроэлектроники в области
изготовления больших интегральных
микросхем. Такая гибридная
интегральная микросхема должна
иметь два вида выводов (рис. 2):
электронные - для соединения с
периферийным электронным
оборудование, включая ЭВМ, и
гидродинамические - для соединения
с периферийными
гидродинамическими устройствами
(другими аналогичными
микросхемами, макрорезервуарами с
питательной средой, макронасосами,
макрооксигенаторами,
макроадсорбционными колонками и т.
д.) (3). Электронная
биогидромеханическая система
должна позволить создать
необходимые условия для запуска in
vitro процесса формирования
функционирующей капиллярной сети
(рис. 3) в желе-
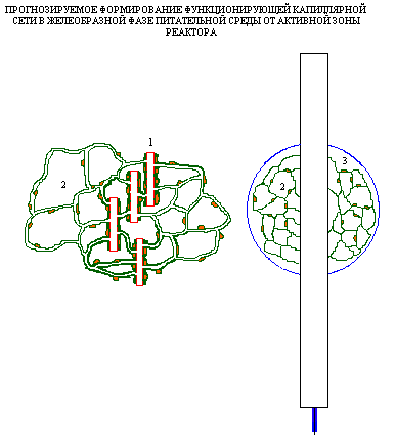
Рис.
3
образной фазе питательной среды (2) от активной зоны сопряжения реактора (1), которая формируется до начала микроциркуляции жидкой фазы питательной среды, путем помещения в нее в специальных условиях культуры эндотелиальных клеток, которые после ряда инженерных манипуляций на клеточном и тканевом уровне должны покрыть снаружи пористые капиллярные микротрубочки сплошным слоем. После начала микроциркуляции эндотелиальные клетки, расположенные на поверхности пористых капиллярных микротрубочек, через поры соответствующего размера в стенке последних будут иметь множество микроконтактов с организованными по желанию экспериментатора микропотоками жидкой фазы питательной среды. Еще до начала микроциркуляции эндотелиальные клетки в гидростатической желеобразной фазе питательной среды должны начать самопроизвольно устанавливать контакты друг с другом, в том числе и между эндотелиоцитами соседних капиллярных микротрубочек, формируя прообраз капиллярной сети, кое-где в эндотелиоцитах должны начать формироваться вакуоли (прообразы внутреннего просвета капилляров). Если пор в капиллярных микротрубочках достаточно много, то, после начала микроциркуляции в последних, от некоторых эндотелиоцитов, закрывающих поры может начаться полноценное формирование канализированного капилляра в просвет которого будет поступать питательная среда из просвета капиллярной микротрубочки через соответствующую пору в стенке последней.
Силами компетентных специалистов из СГМА, СФМЭИ, СГПИ, СОИП, СВА ПВО СВ РФ (г. Смоленск) начаты НИР и ОКР по указанным выше системам реактора. В проекте РФФИ №94-04-13544 автором были выполнены работы по биофизико- и математико-морфологическому моделированию морфологических феноменов в микрососудистых сетях, необходимые для разработки указанной выше гибридной интегральной электронной биогидромеханической микросхемы.
Предложен принципиально новый подход к решению проблемы формирования в клеточной культуре эндотелия in vitro функционирующих саморазвивающихся капиллярных сетей. Полученные результаты соответствуют мировому уровню новизны. Проведены исследования направленные на биофизико- и математико-морфологическое моделирование морфологических процессов в микрососудистых сетях, математическое моделирование микронасосов в предполагаемой гибридной интегральной электронной и биогидромеханической микросхеме на основе пьезокерамической капиллярной микротрубочки.
Исследование выполнено при поддержке Российского фонда фундаментальных исследований
(Проект
РФФИ №96-04-50991).
Кафедра анатомии человека
Смоленская государственная медицинская академия
Поступила в редакцию 15.01.97.