Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 7. -
Вып. 4. - 2008. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTМ
http://www.smolensk.ru/user/sgma/MMORPH/N-20-html/TITL-20.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-20-html/cont.htm
УДК 572
ДИНАМИКА ДИВЕРГЕНЦИИ ВИДОВ В ПОСТНАТАЛЬНОМ ОНТОГЕНЕЗЕ
© 2008 г. Харитонов
В. М.
Работа посвящена сравнительному анализу ростовых процессов черепа ископаемых гоминид в постнатальном онтогенезе, которые могут повлиять на сходство (различие) палеонтологических объектов в процессе классификации.
Ключевые слова: рост, постнатальный онтогенез, череп, гоминиды, дивергенция, таксономия, классификация
Для практики классификации ископаемых и современных гоминид важным представляется вопрос о значении конкретных периодов онтогенеза для дивергенции видов, ибо, классифицируемые палеоантропологом, обьекты могут иметь несходный биологический возраст. Классик приматологии А. Шульц (Schultz,1926) считал возможным лишь возрастание дивергенции признаков видов приматов в позднем онтогенезе. Подобный взгляд на возможную связь дивергентной эволюции и поздней онтогении имеет длительную традицию в зоологии (Познанин, 1963).
М. Ф. Нестурх (1960) считал, что послеутробный период не определяет всех различий, он модифицирует уже намеченные в эмбриональном периоде. Так, пропорции руки (относительная длина плеча, длинная кисть и короткое предплечье) у эмбрионов млекопитающих более сходны чем у взрослых особей. В отношении лицевого скелета приматов подобного взгляда придерживался В. В. Бунак (1960), анализировавший перестройку ростовых процессов, играющую ведущую роль в процессе трансформации черепа. И ранее В. В. Бунак (1959) констатировал, что дивергенция признаков, существующих до рождения, лишь возрастает после него. Постнатальный онтогенез характеризуется определенными сдвигами в интенсивности относительного роста, писал он (Бунак В. В.,1961), в итоге, отрицая влияние распределения относительных скоростей роста (т.е. градиентов роста) поздней онтогении на особенности дефинитивного строения скелета.
Однако, следуя за Я. Я. Рогинским (1977), следует заметить, что наибольшая дивергенция приматов и человека по таким признакам, как величина мозга, характерные пропорции черепа возникает именно после рождения. Он на примере возрастной динамики относительных длин грудных и тазовых конечностей (по отношению к длине туловища) доказал существование сложной и несходной динамики роста у антропоида гориллы и современного человека. Длинорукость гориллы достигается за счет внутриутробного подъема относительной длины, а у человека - длиноногость за счет увеличения относительной длины конечности после рождения. Т.е. динамика дивергенции указанных двух признаков у гоминоидов сложным образом реализуется в пренатальный и постнатальный периоды рождения. Изменения градиентов роста, как формы временной и пространственной организации его, в конечном итоге определяют динамику дивергенции количественных признаков.
В. З. Юровская (1983) справедливо полагала, что направление и скорость изменения признаков могут быть неодинаковыми в разные периоды онтогенеза. Соответственно и дивергенция видов по указанным признакам будет иметь сложный характер. Реализация краниокаудального градиента роста конечностей и их сегментов зависит от сформированного в процессе отбора способа локомоции. Так по некоторым признакам горилла и современный человек равным образом отличны от других приматов (самое короткое предплечье, относительная длина первого пальца кисти).
Рубеж рождения для ряда признаков – время их состояния, отличного от взрослых форм. Вес при рождении у гоминоидов дает ряд, не повторяющий их таксономического положения: современный человек - 5,6 % от взрослого состояния, орангутан - 4,1%, шимпанзе - 4,0%, горилла - 2,6%.
В пренатальном онтогенезе (Жеденов В. И., 1962) относительная величина головы у ранних плодов обезьян больше чем у плода современного человека, на рубеже рождения - нет.
Н. Хайнц (Heintz,1966) уделила большое внимание постнатальному онтогенезу в связи с наблюдаемой значительной дивергенцией ростовых процессов у гоминоидов после рождения, способных формировать таксономически значимые различия в пропорциях черепа.
Обобщая приведенные выше точки зрения, можно сказать, что в пренатальном периоде онтогенеза, на рубеже рождения и в постнатальном онтогенезе можно отыскать примеры такой дивергенции признаком, которая не соответствует классификации дефинитивных форм.
Методом канонического анализа нами исследовалось взаимное положение инфантильных, ювенильных и взрослых форм ископаемых гоминид, объединенные в компьютерную базу данных, в аспекте изучения динамики достижения дефинитивного состояния размера и формы.
Профессор Я. Я. Рогинский (1978) в известной работе разбирал вопрос соответствия онтогенетических дифференцировок филогенетическим. Наша задача скромнее - поиск соответствия поздних онтогенетических перестроек установленным таксономическим взаимоотношениям гоминид.
Исследованные разновозрастные гоминиды относятся к трем основным условным стадиальным группам (архантропы, палеоантропы и неоантропы, включая “архаичных” сапиенсов), представляя четыре одонтологических возраста, выделяемых для приматов А. Шульцем (инфантильный, ювенильный 1, ювенильный 2, взрослый) (Schultz, 1926). Измерительные признаки черепа анализировались в абсолютной форме.
В проведенный анализ вошли: ювенильный синантроп 3; инфантильные палеоантропы Пешь дель Азе, Гибралтар 2, Шатонеф, Баракай; ювенильные палеоантропы Тешик-Таш, Ля Кина, Шипка, Малярно, Заскальная 6; инфантильные "архаичные" сапиенсы Схул 1 и Староселье; инфантильные ископаемые неантропы Пшедмости 2, Пшедмости 6, Младеч 3, Костенки15; ювенильные ископаемые неоантропы Пшедмости 7, Пшедмости 22, Костенки 18, Ниа, Сунгирь 2, Сунгирь 3, Пато, Пшедмости 5.
При постановке задачи исследования мы исходили из того, что канонический анализ может чутко реагировать на изменение в онтогенезе формы полного черепа и его частей. Расстояния между обозначениями гоминид на графиках канонического анализа являются не просто отображением ростового пути, но и динамики изменения формы черепа. Ниже приведены результаты анализа, проведенного указанным выше статистическим методом, до сих пор не применявшимся для исследования ростового пути черепа гоминид. Размещение на одном поле разновозрастных гоминид позволяет визуализировать не только степень их морфологического несходства, но и дивергенцию признаков величины и указателей формы черепа с возрастом.
1. Основанием динамики онтогенетической дивергенции краниометрических признаков являются особенности реализации кефало-каудального градиента роста в пределах черепа гоминид. Анализ обобщенных расстояний между неполовозрелыми и дефинитивными формами гоминид по признакам кальвариума (мозговой и лицевой отделы вместе) показал, что в антропогенезе не наблюдается закономерного изменения различий между неполовозрелыми и взрослыми формами при переходе с одного эволюционного уровня на другой. Ранние и поздние гоминиды различаются в границах фаз постнатального онтогенеза в разных отделах черепа различным образом. Так у ювенильных форм архантропов различие по комплексу признаков кальвариума с дефинитивными формами меньше чем у разновозрастных палеоантропов, "архаичных" сапиенсов и кроманьонцев (т.е. ростовой путь после рождения меньше) (рис.1).
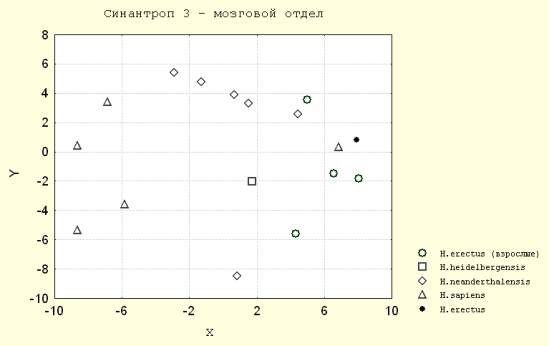
Рис. 1. Синантроп 3(Juvenis) – мозговой отдел
2. Наоборот, анализ результатов по комплексу признаков нижней челюсти выявил преобладание различия инфантильных и дефинитивных форм, ювенильных и дефинитивных форм палеоантропов над различием более поздних гоминид разных стадий онтогенеза (т.е. ростовой путь мандибулы у неандертальцев после рождения больше) (рис.2). Таким образом, п.п. 1 и 2 позволяют предположить, что постнатальный онтогенез у ранних видов рода Нomo характеризуется пролонгированными ростовыми процессами нижнечелюстного отдела лицевого черепа и невыраженным общим приростом кальвариума.
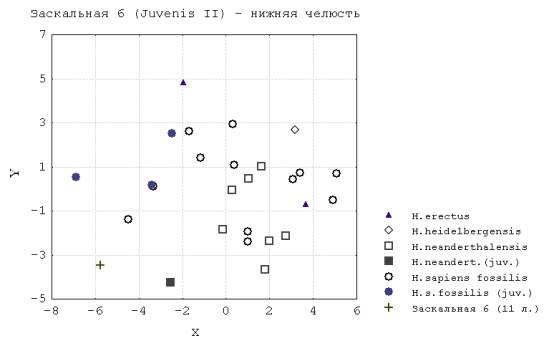
Рис. 2. Заскальная 6 (Juvenis II ) – нижняя челюсть
3. Обратимся к двум традиционным частям черепа. По признакам кальвариума на уровне ювенильного 2 возраста различие разновозрастных архантропов меньше чем между ювенильными и дефинитивными формами палеоантропов и ископаемыми неоантропов. Причем ростовой путь кальвариума у архантропов меньше чем у палеоантропов, а у последних - чем у кроманьонцев. Данное явление объясняется меньшими размерами и сходной формой мозгового черепа у ранних представителей рода Homo различного индивидуального возраста.
Отличие по признакам кальвариума инфантильных,
ювенильных палеоантропов с дефинитивными
формами большее чем у кроманьонцев можно объяснить большими размерами
лицевого отдела у неандертальцев и пролонгацией его роста (рис. 3).
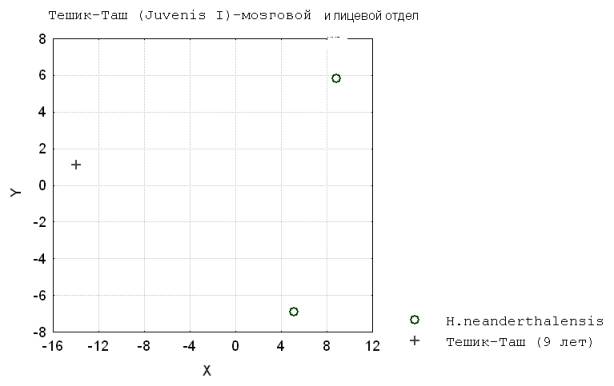
Рис. 3. Тешик-Таш (Juvenis I) – мозговой и лицевой отдел
У ископаемых неоантропов во всех трех одонтологических возрастах зафиксирован высокий уровень отличия с дефинитивными
формами. В этом случаи филогенетический рост мозгового отдела в антропогенезе определил большую величину
ростового пути и интенсивность прироста его в онтогенезе, а также постепенность
оформления взрослых пропорций (рис.4).
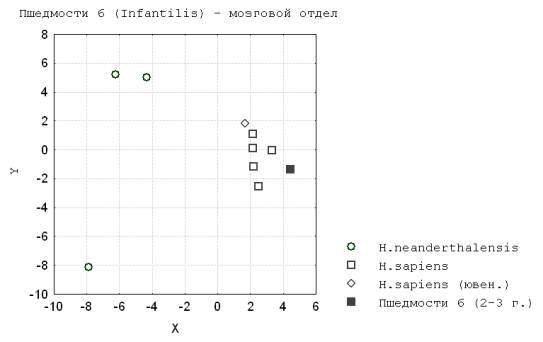
Рис. 4. Пшедмости 6 (Infantilis) – мозговой отдел
4. У палеоантропов высокий уровень отличия неполовозрелых и взрослых форм наблюдается во всех трех одонтологических возрастах по признакам нижней челюсти (рис.5). Это объясняется значительной величиной данного отдела черепа и пролонгированным типом его роста после рождения. Ростовой путь и степень несходства по форме нижней челюсти у разновозрастных палеоантропов превышает наблюдаемый у кроманьонцев.
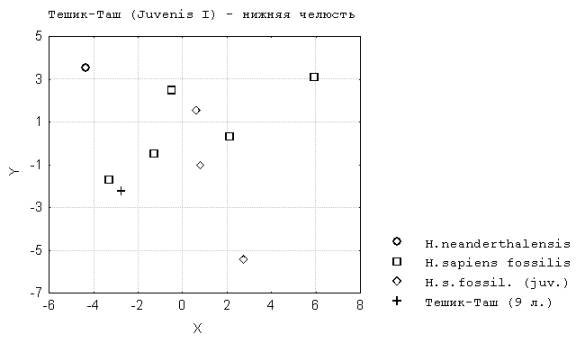
Рис. 5. Тешик-Таш (Juvenis I) – нижняя челюсть
У кроманьонцев различие по комплексу признаков мандибулы в направлении от инфантильного к ювенильным 1 и 2 возрастам уменьшается, нижняя челюсть меньше по размерам чем у других гоминид и, соответственно, по интенсивности роста (рис.6).
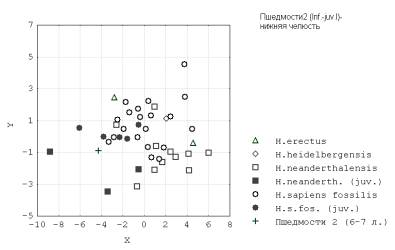
Рис. 6. Пшедмости 2 (Infantilis-juvenis I) – нижняя челюсть
5. Ископаемый инфантильный сапиенс Староселье (Крым) (35 тыс. лет), хранящийся в Музее антропологии МГУ, по степени различия с предполагаемым взрослым состоянием (неандертальским или сапиентным) по гипотетическому направлению роста кальвариума и мандибулы более сходен с палеоантропами (рис.7; рис.8).
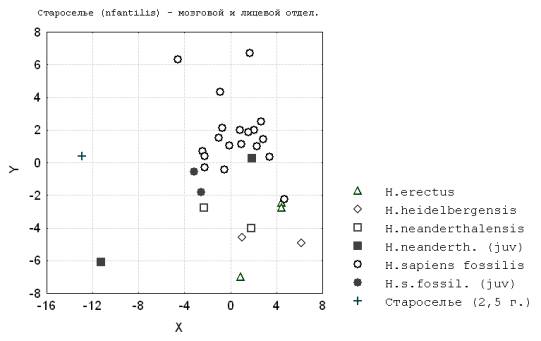
Рис. 7. Староселье (Infantilis) - мозговой и лицевой отдел
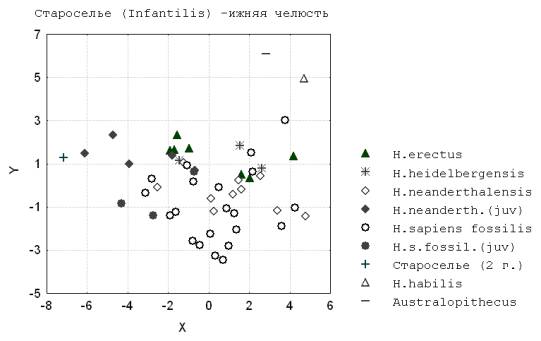
Рис. 8. Староселье (Infantilis)
– нижняя челюсть
6. Рассмотрев все возможные пути возрастного изменения кальвариума у ископаемых гоминид, мы видим, что архантропы почти всегда по величине расстояний между разновозрастными индивидуумами отличны от палеоантропов ( максимальное возрастное отличие их меньше минимального у палеоантропов).
Ископаемые неоантропы также достаточно сильно отличаются от палеоантропов по возрастным потокам значений (минимальные значения различны у первых и вторых) (рис.9).
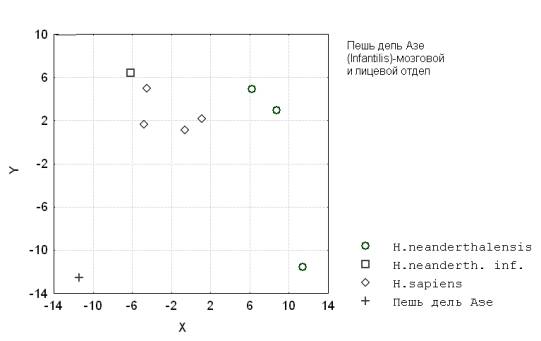
Рис. 9. Пешь дель Азе (Infantilis) - мозговой и лицевой отдел
7. Все наблюдаемые различия у неполовозрелых и взрослых форм ископаемых неоантропов, что любопытно, маркируют ростовой путь на графиках для мандибулы более протяженный чем у других гоминид (рис.10) (продолжительность роста преобладает над приростом нижней челюсти).
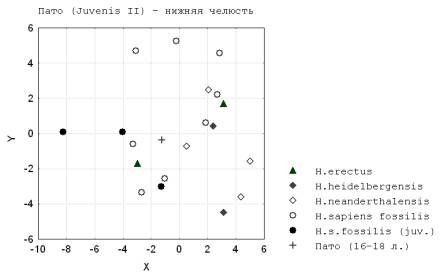
Рис. 10. Пато (Juvenis II)-
нижняя челюсть
8. Данные п.п. 6 и 7 позволяют предположить, что ростовые потоки краниометрических значений в постнатальном онтогенезе имеют классификационное значение.
9. Наконец, зададимся вопросом, как меняется с возрастом масштаб различий ископаемых гоминид разных видов в поздней онтогении?
В работе установлено, что с возрастом от рубежа инфантильного и ювенильного 1 возрастов различия между архантропами и кроманьонцами, между палеоантропами и кроманьонцами по кальвариуму возрастают. По комплексу признаков нижней челюсти отличия палеоантропов и кроманьонцев после рождения уменьшаются.
Приведенное наблюдение подтверждает предположение Я. Я. Рогинского: морфологическое отличие различных видов гоминид обусловлено конкретной модификацией кранио-каудального градиента роста в пределах черепа.
"Архаичный" инфантильный сапиенс Схул I (Израиль) (90 тыс. лет), считавшийся раннее неандертальцем, демонстрирует с инфантильными формами данного вида отличия по кальвариуму меньшие чем те, что существуют между взрослыми палеоантропами и кроманьонцами. У ископаемого сапиенса Староселье (Крым) в младенческом состоянии с инфантильными кроманьонцами отличия по признакам кальвариума больше чем между взрослыми кроманьонцами и неандертальцами, а по мандибуле - меньше. Налицо переходный характер данного гоминида по ростовым характеристикам.
10. Дефинитивные комплексы признаков черепа и нижней челюсти у видов H.erectus, H.neanderthalensis и H.sapiens, что видно на графиках канонического анализа, имеют разную динамику становления, что необходимо учитывать при сопоставлении гоминид заметно различных по биологическому возрасту.
11. Большой интерес для антропологов представляют кроманьонские дети из верхнепалеолитического памятника Сунгирь (окрестности г. Владимира) (около 24 тыс.лет). Анализ графиков показал, что по комплексу признаков мозгового и лицевого черепа Сунгирь 2 (13 лет) и Сунгирь 3 (10 лет) сильно отличаются от неандертальского ребенка Тешик-Таш (9 лет). Что касается роста мандибулы, то предполагаемый рост ее до значений взрослых кроманьонцев у разновозрастных детей из Сунгиря между собой не различается, что, может объясняться достаточно большой интенсивностью прироста ее у младшего ребенка - проявление морфологического своеобразия.
12. О росте нижней челюсти неполовозрелых неандертальцев, исследованных с помощью канонического анализа, можно сказать следующее. Инфантильные формы (2,5 - 5 лет) имели значительный ростовой путь до взрослого состояния. Нижняя челюсть неандертальцев интенсивно прирастала после рождения.
Ювенильному неандертальцу Шипка (9 лет) предстоял ростовой путь значительно меньше. На графиках она расположена в области значений взрослых форм неандертальцев. У подростка - неандертальца Тешик-Таш того же возраста, дистанция с взрослыми неандертальцами больше, что объясняется, видимо, большей величиной мандибулы. Ростовой путь Тешик-Таша (9-10 лет) и Заскальной 6 (11 лет) значителен на фоне разброса значений разновозрастных кроманьонцев.
13. Динамика изменения мозгового и лицевого отдела черепа у палеоантропов после рождения выглядит следующим образом. Кальвариум инфантильного неандертальца (2,5 года) имеет значительный ростовой путь до взрослого состояния , а у ювенильного (8 лет) он значительно меньше. При этом, подросток Тешик-Таш (9 лет) имеет значительную дистанцию со взрослыми формами неандертальцев, что объясняется значительными процессами роста и формообразования лицевого отдела, характерными для него.
Таким образом, на материале изученных неандертальцев мы не видим равномерного снижения различия между инфантильными, ювенильными и дефинитивными формами в постнатальном онтогенезе по признакам разных отделов черепа.
14. Ростовые и формообразующие процессы нижней челюсти неполовозрелых форм кроманьонцев, изученные с помощью канонического анализа выглядят следующим образом. Инфантильный кроманьонец (5,5 лет) имеет значительный ростовой путь до взрослых форм своего вида. Он не меньше того, что характерен для инфантильных форм неандертальцев. Ростовой путь мандибулы ювенильных форм кроманьонцев меньше. Таким образом, ростовой путь мандибулы меняется с возрастом у кроманьонцев закономерно.
15. В границах постнатального онтогенеза кальвариум у неполовозрелых форм кроманьонцев по данным канонического анализа менялся следующим образом. У инфантильных форм (2,5 - 6-7 лет) минимальное различие со взрослыми формами невелико. У изученных ювенильных форм (9,5-17 лет) оно может достаточно выраженным или небольшим. Таким образом, не так очевидно возрастное закономерное изменение отличия разновозрастных форм по кальвариуму, что может быть объяснено верхнепалеолитическим краниологическим полиморфизмом.
Приведенный в работе анализ разновозрастных ископаемых гоминид детализирует существующие точки зрения на связь дивергенции видов гоминид и ростовых процессов поздней онтогении. Результаты его следует учитывать при введении в классификацию неполовозрелых индивидуумов гоминид (даже поздних), так как существуют особенности в возрастной морфологии черепа, которые могут несколько модифицировать наблюдаемые различия между гоминидами в конкретных предефинитивных возрастах. Ростовой процесс в постнатальном онтогенезе не ведет прямолинейно к достижению дефинитивного размера и взрослой формы, что исключительно важно для объективной таксономии.
ЛИТЕРАТУРА
1. Бунак В. В. Череп человека и стадии его формирования у
ископаемых людей и современных рас.
//Труды Ин-та этнограф.,н.с., 1959, Т.XLIX.
2. Бунак В. В. Закономерности относительного роста как
основного фактора формообразования в позднем (постэмбриональном) онтогенезе.
//Архив анатомии, гистологии и эмбриологии, 1961, № 2.
3. Жеденов В. И. Сравнительная анатомия приматов. - М.,
1962.
4. Нестурх М. Ф. Приматология и антропогенез. - М., 1960.
5. Познанин Л. П. Эколого-морфологическое изучение
древесных птиц и вопросы соотношения
таксономической и экологической классификации птиц. - Киев, 1963.
6. Рогинский Я. Я. Проблемы антропогенеза. - М.,1977.
7. Юровская В. З. Сравнение онтогенеза человека и
приматов. //Морфология человека. - М., 1983.
8.
Heitz N. Application de l`etude de la croissance du crane a celle De la
phyloogenese//Acta Fac/ rerum natur. Univ.Comenianae.Anthrop., 1966, 10, N 8.
9. Schultz A. Fetal
growth of man and other primates. N.-Y.,1926.
Dynamics
of species´ divergence in postnatal ontogeny.
Kharitonov
V. M.
Paper is dedicated to
comparative analysis of Fossil Hominids´ skull growth processes in
postnatal ontogeny that may cause similarity (distinction) of paleoanthropologic
objects in the course of classification.
Key words: growth processes,
postnatal ontogeny, skull, classification
Лаборатория антропогенеза
Научно-исследовательский институт и Музей антропологии
МГУ имени М .В.
Ломоносова
Поступила в редакцию 28.10.2008.