Математическая морфология.
Электронный математический и медико-биологический
журнал. - Т. 8. -
Вып. 3. - 2009. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-23-html/TITL-23.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-23-html/cont.htm
УДК 58.037
АДАПТАЦИЯ ДЕРЕВЬЕВ К АНОМАЛЬНЫМ ФИЗИЧЕСКИМ ФАКТОРАМ
Ó
2009 г. Холманский А. С.
Установлено наличие специфических
реакций на действие внешних физических факторов у хвойных и лиственных пород
деревьев. Для первых характерен аномальный спиротропизм (закручивание структуры
древесины вокруг оси дерева), а для вторых – аномальный электротропизм,
конкурирующий с геотропизмом при определенных условиях. Данные различия в
механизмах адаптации деревьев к внешним физическим факторам связали с отличиями
электрофизических и хиральных свойств жидкой среды и морфологии лиственных и
хвойных пород. Используя гипотезу о биогенности солнечного нейтрино, предложили
возможный механизм его действия на деревья в качестве внешнего хирального
фактора ответственного за явление спиротропизма.
Ключевые слова: деревья, электротропизм, спиротропизм, аномальные поля, нейтрино.
1. ВВЕДЕНИЕ
Адаптационные изменения структуры и формы живых организмов формально подчиняются принципу Кюри, устанавливающему общее правило изменения симметрии упорядоченных структур под воздействием факторов внешней среды (механических, электромагнитных). К структурной самоорганизации молекул в живых системах применимы принципы организации жидких кристаллов, поэтому их динамическую стереометрию вполне можно рассматривать как производную стереохимии органических соединений и симметрии внешних физических факторов. Процесс согласования элементов симметрии внутренних и внешних факторов лежит в основе механизма адаптации растений.
При изучении феногенеза растений уже выявлена специфика реакций метаболизма на действия гравитации, электрического поля, света, температуры, воды, химических веществ и других факторов, обусловливающих то или иное изменение во внутренней структуре и внешней форме растения (тропизм). Однако до сих пор до конца не ясен физико-химический механизм тропической реакции растения на известные физические факторы и совсем не известна природа фактора ответственного за скручивание древесины дерева. Данное явление в слабой форме присуще многим деревьям, кроме того, часто встречается значительное закручивание стволов деревьев разных пород с направлением закрутки, как правым, так и левым по отношению к направлению их роста [1]. Такое изменение внешней формы и внутренней структуры дерева можно назвать спиротропизмом.
В соответствие с принципом Кюри внешний фактор, обусловливающий спиротропизм деревьев, должен иметь спиральную симметрию и переменный знак, то есть быть хиральным (J-фактор [2]). В общем случае чувствительность живой системы к J-фактору будет определяться наличием в ее структуре атомно-молекулярных элементов с хиральной тетраэдрической метрикой. Роль J-фактора, в принципе, могут играть вихревое магнитное поле, циркулярно поляризованный свет и нейтрино.
Различают внутреннее и наружное закручивание деревьев. Последнее подразумевает закручивание ствола как целого при отсутствии заметного закручивания текстуры древесины. Примерами чисто наружного закручивания может служить обвивание виноградной лозой опоры или закручивание сосен в правые спирали на Куршской косе. Данные явления, также как причудливые формы сосен в скалистой местности или изломы карельских берез следует отнести к аномалиям геотропизма, обусловленным внешними факторами иной, чем J-фактор природы.
Считают [1], что причиной наружного закручивания могут быть вирусные заболевания, наследственность или особенности роста. Иногда этот признак начинает передаваться по наследству и становится характерным для всего вида. Наружное закручивание или изгибание наблюдается особенно часто у деревьев, растущих в специфических условиях – в тропиках, у верхней границы высокогорных лесов, на каменистых утесах, в равнинных пустынях, в песчаных прибрежных районах (Куршская коса) или на северных широтах (карельская береза). Для подобных местностей характерны геофизические аномалии (гравитационные или электромагнитные), переменные ветры, резкие и значительные перепады влажности и температуры, жара и засуха.
В полной мере спиротропизму отвечает внутреннее закручивание, которое проявляется на симметрии текстуры древесины. У хвойных деревьев оно обычно скрыто корой, но у некоторых лиственных (например, каштан [1]) и этот слой дерева также участвует в закручивании. Направление закручивания ствола может быть различным в разных слоях древесины и на разных высотах ствола. У некоторых культурных видов, а также у многих тропических деревьев направление закручивания меняется через довольно частые и равные интервалы. В этих случаях в последовательных слоях древесины по радиусу ствола чередуются правосторонняя и левосторонняя спирали, причем каждый слой может включать несколько годовых колец. Варьируя, например, с помощью температуры и влажности водоснабжение дерева многократно вызывали правое и левое закручивание ствола у молодых караибских сосен [1]. Воздействие других неблагоприятных факторов среды – таких, как интенсивность света, ионизирующая радиация, угнетение другими деревьями и длина дня – не вызывало закручивания. До сих пор нет объяснений ни механизма закручивания, ни того, почему растение сохраняет спиральное направление роста.
Следует отметить, что спиротропизм деревьев и ресурс функциональной асимметрии мозга человека [3] могут быть обусловлены действием биогенных квантов одной и той же космической природы (J-фактор). В работах [2 – 4] для обоснования нейтринной природы J-фактора предложен механизм биогенизации квантов нейтринной энергии сплошными кристаллическими и жидкими средами в составе геосфер и живых организмов. В основе данного механизма лежат кооперативные эффекты пространственной организации в атомных и молекулярно-клеточных подсистемах, объединяющих большое число однородных элементов. В результате этих эффектов возмущающий характер действия J-фактора усиливается до уровня энергетики метаболических реакций и приобретает роль хирального кинетического фактора.
Очевидно, что для прямой регистрации внешнего J-фактора спиротропизма нужны методы, чувствительность, которых на порядки превышает чувствительность, например, таких методов, как электро- или магнитоэнцефалография. Биогенность, хиральность и вездесущность квантов солнечного и галактического нейтрино (антинейтрино) позволяют принять его за глобальный фактор асимметрии, отвечающий в частности за генезис спиротропизма и полового диморфизма у растений и животных.
В настоящей работе изучили зависимость спиротропизма и некоторых аномалий геотропизма от породы деревьев и условий внешней среды с целью выявления физико-химических особенностей в адаптации живых систем, свидетельствующих о чувствительности метаболизма растений к J-фактору.
2. МЕТОДИКА И РЕЗУЛЬТАТЫ
Очевидно, что механизм адаптации должен зависеть от породы дерева и условий их произрастания. Это, прежде всего, относится к хвойным и лиственным деревьям, произрастающим в северном полушарии и имеющим существенные отличия в микроструктуре древесины и свойствах ее жидкой среды. Типичными их представителями являются сосна (обыкновенная и кедровая), ель, лиственница, береза и дуб. Особенности тропизмов и закручивание этих деревьев изучили на крутых склонах (наклон ~45о) естественного или искусственного происхождения, а также в местах с аномальной геофизикой. Оказалось, что на склонах сосны и ели строго сохраняют радиальное направление (по радиусу Земли), тогда как березы достаточно часто (~30% деревьев) вблизи от земли ориентированы почти ортогонально к поверхности склона, а затем изгибаются и растут в радиальном направлении (Рис 1 b, с).
Электрофизические свойства древесины и почвы изучали путем измерения разности потенциалов (V) между деревьями различной породы и землей с помощью портативного тестера (MAS838), используя в качестве контактных электродов железные гвозди (длиной 30, 70 и 100 мм) и полоски из нержавеющей стали (2×10×150 мм). Данная электрическая система представляет собой гальванический элемент: {Fe|почва||мембрана||древесина|Fe} с одинаковыми электродами, различными электролитами, разделенными полупроницаемой мембраной гидрофобной кутикулы корневой системы дерева. В древесину электрод углубляли на 1,5 – 2 см вплоть до твердых слоев ксилемы, а в землю – на всю длину электрода. При использовании в качестве электродов гвоздей фиксировали разность электродных потенциалов и ток (I), значения которых оставались стабильными в течение часа. На электроде, контактирующем с жидкой средой древесины возникал отрицательный потенциал, а на электроде, погруженном в землю – положительный, соответственно ток электронов был направлен от первого электрода ко второму. Величина V (береза, кактус) уменьшалась в 2-3 раза, если измерения проводили через сутки и более при неизменном положении электродов. При этом на части гвоздя находящейся в березе и, особенно в кактусе образовывался слой ржавчины. Результаты измерений V «земля-дерево» и I для деревьев различного вида и диаметра, а также для комнатного кактуса представлены в таблицах 1, 2.
При замене гвоздей на полоски из нержавеющей стали фиксировали разность потенциалов того же знака, но она снижалась от ~0,3 V почти до нуля за время порядка 15 мин. Начальный ток в такой системе был существенно меньше (0,01 мА), чем в элементе с контактами из гвоздей, и он снижался до нуля за время порядка ~3 секунд.
Таблица 1
Разность потенциалов «земля-растение» для различных видов растений
|
N |
Растение, диаметр ствола (см) |
V (B) |
|
1 |
Сосна, 10 ÷ 40 |
0,16 ÷ 0,30 |
|
2 |
Лиственница, 15 ÷ 30 |
0,11 ÷ 0,21 |
|
2 |
Ель,15 ÷ 25 |
0,12 ÷ 0,22 |
|
3 |
Береза, 10 ÷ 30 |
0,18 ÷ 0,50 |
|
4 |
Дуб, 20 ÷ 70 |
0,18 ÷ 0,32 |
|
5 |
Липа, 30 |
0,25 |
|
6 |
Черемуха, 20 |
0,27 |
|
7 |
Слива Живая, 12 Мертвая, 12 |
0,40 0,34 |
|
8 |
Ольха, сухая, 20 |
0 |
|
9 |
Кактус комнатный, 2 |
0,13 |
Значение V в случае березы оставалось практически неизменным при изменении высоты гвоздя в дереве и расстояния между березой гвоздем в земле в пределах 10 – 100 см (Таблица 2). Для мертвых (лишенных зелени, с сухими ветвями), но еще сохраняющих влагу деревьев величина V была, как правило, незначительно ниже, чем для живых (см. слива в Таблице 1).
Таблица 2
Зависимость разности потенциалов и тока
от расстояния между березой и электродом в земле
|
N |
L (см) |
V (B) |
I (мкA) |
|
1 2 3 4 5 |
10 20 30 50 100 |
0,37 0,37 0,36 0,36 0,36 |
34 36 31 34 31 |
У полностью высохших деревьев величина V равнялась нулю. У пня двухгодичной давности величина V возрастала в 1,5 - 2 раза при перемещении гвоздя от центра пня к периферийным слоям древесины, лежащим на расстоянии ~2 см от поверхности дерева, достигая значений близких к V для живого дерева (~0,3V). Потенциал гвоздя в чурке свежесрубленной березы лежащей на боку на влажной земле возрастал вдвое при переворачивании ее на попа (от 0,2 до 0,4 В).



a
b c
Рис 1. Сосна обыкновенная с правовинтовой закруткой – a (Красногорск, Московская обл); сосны кедровые на склоне горы в Саянах – b; береза на склоне берега канала Москва-Волга в районе г. Долгопрудного – с.
Закрученность стволов изучали в основном по сухим деревьям, чьи стволы частично не имели коры. Спиральность или закрученность древесины (J±) оценивали по величине скоса трещины ствола (Δ в сантиметрах) на погонном метре его высоты (J± = Δ/100). При этом скос против часовой стрелки отвечал правовинтовой закрученности (J+), а по часовой – левовинтовой (J–). Наблюдения показали, что у сосны и ели (Рис 1) в некоторых местах около ~5% деревьев имеют правовинтовую закрученность J+ ~ 0,1 ÷ 0,3. У березы и дуба закрученность, как правило, отсутствует. Стальная иголка, подвешенная на тонкой нити у основания закрученного дерева (сосна, ель) оставалась в покое, ориентируясь параллельно стрелке компаса, что указывает на отсутствие в данном месте переменной составляющей земного магнитного поля.
Очевидно, что самым чувствительным к J-фактору элементом древесины будет ее жидкая среда, содержащая оптически активные метаболиты. К последним у лиственных и хвойных деревьев относятся, прежде всего, аминокислоты и сахароза. Кроме этого, у хвойных деревьев существенный вклад в спиротропизм может вносить и живица, в составе которой содержаться оптически активные скипидар и смоляные кислоты. Для выявления сезонных вариаций хиральности жидкой среды деревьев в работе провели мониторинг оптической активности физиологического 10% раствора декстрана (реополиглюкина) и скипидара в период весеннего пробуждения деревьев, вплоть до появления первой зелени (~24.04.09 в Долгопрудном). Использовали круговой поляриметр СМ-3 с кюветами 100 и 200 мм, измерения проводили при комнатной температуре (24,5 ± 0,5оС). Ход зависимости оптической активности, характеризуемой величиной угла поворота плоскости поляризации света (α), представлен на Рис 2.
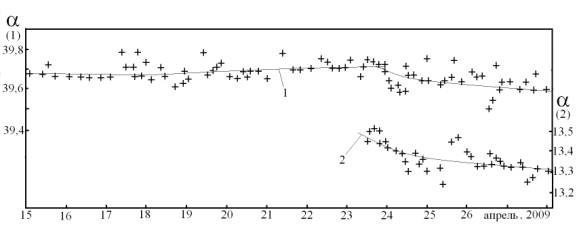
Рис 2. Мониторинг оптической активности физраствора декстрана (1) (кювета 200 мм) и скипидара (2) (кювета 100 мм). Крестики – экспериментальные точки, линия – их среднее значение.
Для выяснения возможности приобретения потоком теплового излучения спиральной анизотропии (хиральности) при прохождении через оптически активную среду изучили влияние на процесс плавления кристаллов сахара-рафинада подложек из различных материалов с аморфной и кристаллической структурой (Таблица 3). Сравнили также зависимость температуры плавления (Тпл) от время суток при нагреве микрокристаллов сахара на алюминиевой болванке (диаметр 3 см) и на лежащей на ней оптически активной пластины сапфира.
Болванку нагревали путем пропускания тока по нихромовой спирали, намотанной на болванку, и ее температуру измеряли с точностью до 0,1оС с помощью ртутного термометра, углубленного в болванку. Процесс плавления микрокристаллов сахара различного размера имел температурный диапазон шириной в ~2оС. В Таблице 3 и на Рис 3 приведены значения Тпл, отвечающие средней температуре диапазона. Скорость нагрева составляла ~1о/мин. За состоянием образцов наблюдали с помощью микроскопа МБС-1 (40×). Измерения проводили ежедневно с 16.07.08 по 06.08.08. Каждая точка на графике (Рис 3) отвечает усредненной величине 3 – 5 измерений Тпл в указанное время и в разные дни.
Таблица 3
Температура плавления кристаллов сахара на различных подложках
|
№ |
Материал пластины подложки |
k* Вт/(м К) |
Толщина пластины (мм) |
Температура плавления (оС) |
|
1 |
Поверхность болванки (Al) |
230(0,026)** |
– |
172,0 |
|
2 |
Силикатное стекло (аморфное, Na2OCaO 6SiO2) |
1,15 |
0,25 |
174,3 |
|
3 |
Оптически активный Сапфир (кристалл, Al2O3) |
35 |
0,32 |
174,2 |
|
4 |
Кристалл кремния (Si) |
150 |
0,39 |
173,0 |
|
5 |
Оптически активный кварц (кристалл, SiO2) |
1,1 |
0,42 |
174,4 |
*) k – коэффициент теплопроводности; **) kAl (kвоздух)
Из данных приведенных в Таблице 3 и на Рис 3 следует, что Тпл практически не зависит от физико-химических свойств подложки и времени суток. Исходя из этого, можно заключить, что, либо хирализация теплового излучения отсутствует, либо процесс плавления кристаллической структуры сахара не чувствителен к спиральной анизотропии потока теплового излучения (ИК-фотоны). Величина разницы в Тпл сахара на подложках и на болванке определяется коэффициентом теплопроводности материала подложки. Свой вклад в нее вносят и тепловые потери в воздушном зазоре между подложками и болванкой.
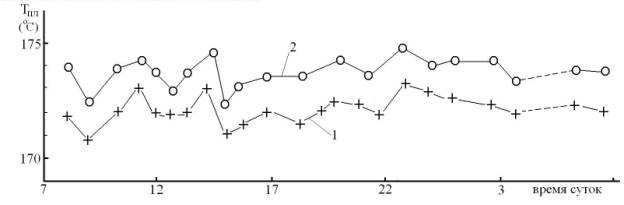
Рис 3. Суточный ход зависимости температуры плавления (Тпл) кристаллов сахара на поверхности алюминиевой болванки – 1 и на пластинке из сапфира, лежащей на болванке – 2.
3.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
3.1. ЭЛЕКТРОФИЗИКА ГЕОТРОПИЗМА
В основе тропизма растений лежит физический механизм усиления потоков энергии, метаболитов и гормонов роста к участкам, молекулярно-клеточная структура и жидкая среда которых испытывает действие того или иного физического фактора. Восприниматься эти действия могут всеми структурами растения, но за тропическую реакцию будут отвечать главным образом живые ткани камбия, то есть цилиндрической латеральной меристемы, локализованной между ксилемой и флоэмой ствола, ветвей и корней. Именно в камбии образуются новые клетки ксилемы и флоэмы ежегодно в течение весны и лета, что и приводит к удлинению и утолщению ствола, ветвей и главных корней. Эта активность камбия регулируется фитогормонами, которые синтезируются в апексе и перемещаются вниз по флоэме.
Предполагают, что для каждого внешнего фактора в растении имеется своя система рецепции, итоговой реакцией которой оказывается соответствующая деформация градиентов потоков заряженных метаболитов через клеточные мембраны. Силу тяжести, например, в растительных клетках воспринимают специальные рецепторы (тяжелые частицы, крахмальные зерна). Полагают [5], что избыточное давление рецепторов на нижние клеточные мембраны приводит к сдвигам в их проводимости и поляризации клеток и органа в целом. На анизотропию межклеточного электрического поля реагирует механизм транспортировки отрицательно заряженного гормона роста (ауксина) – его содержание возрастает у положительно заряженной нижней части клеточного образования. Ткани, обогащенные ауксином способны притягивать питательные вещества, что и обеспечивает ускорение роста именно в нижней части. При этом вертикально стоящий стебель растет вверх, а горизонтально расположенный – изгибается кверху. В корнях механизм геотропизма обратный, что связывают с участием в нем другого фитогормона – абсцизовой кислоты [5].
Таким образом, в основе механизма чувствительности метаболизма дерева к тому или иному внешнему фактору лежат электрическая поляризация внутренней и межклеточной жидкой среды древесины. Очевидно, что электрические поля, связанные с поляризацией жидкой среды зависят от диэлектрической постоянной и концентрации заряженных метаболитов в соке или живице, а также от величины напряженности и направления внешнего электрического поля. Причем интенсивность этих полей должна быть существенно больше стационарных электротонических полей, существующих между различными частями растительного организма [5]. Действительно, установлено, что интенсивность фотосинтеза в листьях возрастает при подачи на растение отрицательного потенциала внешнего электрического поля (~103 В), тогда как при подачи положительного потенциала интенсивность фотосинтеза снижалась [6]. Отрицательный потенциал, очевидно, интенсифицировал транспортировку к листьям метаболитов с положительным зарядом (катионы калия, кальция), ускоряя одновременно нисходящий транспорт отрицательно заряженных продуктов фотосинтеза (ауксина).
Наибольшее содержание воды и живицы достигается в слоях камбия и заболони (до 80-90 % масс [7]), поэтому метаболизм этих тканей будет наиболее чувствителен к эффектам поляризации во внешнем электрическом поле. Действительно, максимальная величина V наблюдается при глубине погружения гвоздя в ткань древесины сравнимой с глубиной залегания камбия. Характер зависимости V и I во внешней цепи гальванического элемента от материала электрода и времени позволяет заключить, что в основе механизма генерации V в элементе с электродами из гвоздей лежит процесс образования равновесного двойного слоя электрических зарядов на границе контакта Fe|электролит. Положительный слой формируется из гидратированных катионов железа, выходящих из гвоздя, а отрицательный – из избыточных электронов в электроде.
Жидкие среды дерева и почвы в силу неразрывности их водной фазы будут составлять единое целое, и скорость распространения фронта ее поляризации будет близка к скорости распространения электрического поля в среде с соответствующей диэлектрической постоянной. В полупроницаемой и поляризованной мембране корневой системы работают активные механизмы транспорта воды и минеральных солей, благодаря которым содержание воды и ионов в ксилемном соке дерева повышается на один, два порядка по сравнению с почвой [5]. Именно поэтому интенсивность процесса гидратации катионов железа и плотность заряда в двойном слое на электроде в дереве оказывается больше, чем на электроде в почве, что и обусловливает направление тока электронов в элементе. Уменьшение равновесной плотности электронов в электроде, погруженном в дерево при протекании тока по внешней цепи элемента восполняется выходом и гидратацией новых катионов железа в количестве необходимом для восстановления зарядового равновесия в двойном слое на обоих электродах.
Окисление поверхности гвоздя снижает интенсивность процесса гидратации катионов железа и плотность зарядов в двойном слое уменьшается, что и приводит к неизбежному снижению величины V со временем. Генерация V в элементе с электродами из нержавеющей стали может идти по такому же механизму, но с участием примесных катионов металлов на поверхности электродов. При этом отрицательных зарядов в электроде практически не образуется, на что и указывает низкий ток в таком элементе и быстрая кинетика его спада. Не исключено также, что потенциал и отрицательный заряд на электроде из нержавеющей стали образуются в процессе разрушения ткани дерева [6].
Из этих данных и того, что значения V и тока в элементе становятся максимальными сразу в момент замыкания цепи и не зависят от длины (сопротивления) «древесного» и «земляного» участков цепи следует, что в обоих электролитах тока не возникает, но происходит поляризация их жидкой фазы. Для исключения влияния электротонических токов отметим, что их величина в растениях в сотни раз меньше величин I (в корневых волосках – порядка 0,1 – 0,4 мкА). Кроме того, скорость движения зарядов, отвечающих за эти токи будет лимитирована скоростью распространения потенциала действия, величина которой у растений составляет 0,08 – 0,5 см/с [5].
Известно [8], что поверхность земли имеет отрицательный заряд, которому отвечает электрическое поле с вектором напряженности ортогональным поверхности земли. Его величина составляет в среднем 150 В/м, однако в некоторых местах и при неустойчивых погодных условиях она может возрастать до ~1000 В/м [6]. Как отмечалось выше, поляризация жидкой среды растения во внешнем электрическом поле и ее влияние на метаболические процессы будет тем значимей, чем выше ее диэлектрическая постоянная. У березы ткань камбия насыщена пасокой, являющейся слабым электролитом с диэлектрической постоянной, близкой к воде (ε ~ 80). У сосны в поверхностных тканях древесины (камбий, заболонь) жидкая среда включает практически равные количества воды и живицы. Основу живицы составляет скипидар, имеющий ε ~ 2. Можно полагать, что средняя величина ε жидкой среды камбия хвойных деревьев будет существенно ниже, чем у лиственных. Поэтому поляризация во внешнем электрическом поле и ее влияние на метаболизм у лиственных деревьев (березы, дуба) будет на порядок сильнее, чем у хвойных пород (сосна, ель). Данный вывод согласуется с тем фактом, что величины V у лиственных пород, как правило, выше, чем у хвойных (Таблица 1).
Характерное искривление ствола березы в некоторых местах на склонах (Рис 1 с) можно связать с электротропизмом, который на малых высотах в местах с аномально высокими значениями напряженности электрического поля земли превалирует над геотропизмом, в силу чего ствол березы ориентируется по направлению силовых линий электрического поля, то есть перпендикулярно поверхности земли. Поскольку кулоновская сила уменьшается с высотой по квадратичному закону, а сила тяжести практически постоянна, то конкуренция электротропизма с геотропизмом возможна лишь на небольших высотах. Данная конкуренция предполагает, что в механизмах геотропизма и электротропизма задействован один и тот же рецептор, реагирующий одновременно на действие силы тяжести и кулоновской силы. Причем восстановление радиального направления роста ствола березы должно происходить на высотах, на которых действие этих сил на клеточный рецептор сравнивается.
Для реализации механизмов гео- и электротропизма потенциальная энергия соответствующего рецептора в гравитационном (Ug) или электрическом (Uq) поле должна превышать его тепловую энергию (kT при 300оС ~4 10–21 Дж). Исходя из этого, можно оценить характерные параметры частицы-рецептора силы тяжести и аномальную величину напряженности электрического поля земли, при которой электротропизм конкурирует с геотропизмом. Порядок пороговых величин потенциальных энергий тяжелой или заряженной частицы в пределах клетки высотой h ~ 10 мкм можно оценить из соотношений:
Uq ~ Ug ≥ kT,
используя следующие выражения:
Uq = Eqh и Ug = ΔρVgh ~ 4ΔρR3gh,
где Δρ есть разница плотностей зерна крахмала и воды – (1,6 – 1,0) 103 кг/м3, а q есть положительный единичный заряд – 1,6 10-19 Кл, адсорбированный частицей-рецептором; g = 10 м/с2. Для зерна крахмала, играющего роль рецептора силы тяжести и имеющего форму шарика радиуса R, его диаметр составит ~0,6 мкм, а величина Е ~ 2 103 В/м, что хорошо согласуется с размерами зерен крахмала в растительных клетках и аномальными величинами напряженности электрического поля земли [6].
Нечувствительность деревьев хвойных пород к электрическому полю земли можно связать с низкой поляризуемостью их жидкой среды, а также с возможным отсутствием заряда на частице-рецепторе силы тяжести. Поэтому ориентация этих деревьев на любом склоне определяется только геотропизмом, и они растут строго по радиусу Земли (Рис 1b). Из электростатики следует, что электрическое поле поверхностных зарядов не проникает вглубь земли, поэтому ориентация корней у хвойных и лиственных пород на склонах определяется только геотропизмом.
3.2. СТРУКТУРНЫЙ ФАКТОР СПИРОТРОПИЗМА
В основе механизма спиротропизма деревьев, очевидно, лежит чувствительность к J-фактору электродинамической постоянной жидких сред камбия (пасоки, живицы), а также механизма движения хиральных метаболитов в сосудах ксилемы и флоэмы. Действие J-фактора на хиральную среду можно проиллюстрировать обратным эффектом Фарадея, в котором цирукулярно поляризованный свет порождает в магнитном поле вихревую составляющую. А в случае потока заряженных метаболитов закручивающие действие на них J-фактора можно сравнить с действием силы Лоренца на двигающийся в магнитном поле заряд. Абсолютная скорость передвижения воды по ксилеме для лиственных пород равна в среднем 0,2 м/час, а для хвойных – 0,05 м/час. Скорость потока веществ по флоэме составляет 0,2 - 1 м/час и ~90 % в нем приходится на сахарозу. Поскольку она практически не участвует в метаболизме растения, ее можно считать носителем фитогормонов, аналогичным воде в восходящем потоке метаболитов.
Известно [5], что ключевую роль в метаболизме растения играет калий, содержание которого в тканях составляет в среднем 0,5 – 1,2 % от сухой массы, а в клетках уровень калия в 100-1000 раз превышает его уровень во внешней среде. Калий необходим для поглощения и транспорта воды по растению и сахарозы по флоэме. Считают, что корневое давление на ¾ обусловлено присутствием в пасоке калия. Калий поглощается в виде катиона и образует лишь слабые связи с различными соединениями в клетке. Ключевая роль катионов калия в работе всех транспортных систем дерева обусловлена тем, что в силу отрицательности энергии его гидратации, молекулы воды в водных растворах калия имеют большую подвижность, чем в чистой воде. С другой стороны специфика взаимодействия катионов калия с водой способствует увеличению его собственной подвижности в гетерогенных средах с амфифильными свойствами. Очевидно, что такие свойства имеют пектиновые каналы в кутикуле, через которые в корневую систему поступают вода, калий и другие вещества из почвы.
Отметим, что пектиновые каналы наряду с высокой адсорбционной способностью могут иметь свойственную полисахаридам оптическую анизотропию. В дополнение к этому из корневых волосков периодически выделяется в почву содержащие сахара выделения (слизь). Считают [5], что слизь увлажняет почву, это способствует диффузии веществ из нее к корню и облегчает движение растущего корня в почве.
Таким образом, в сплошных потоках вещества в самом дереве и в его корневой системе присутствуют хиральные и заряженные метаболиты. Это позволяет предполагать возможность действия на них J-фактора по механизмам аналогичным эффектам Фарадея и Лоренца. О наличии чувствительности к J-фактору жидкой среды древесины можно судить по суточным и сезонным изменениям угла вращения раствора декстрана и скипидара, моделирующих содержащие сахара жидкие и гелеобразные среды дерева, а также живицу (Рис 2). Снижение их уровня хиральности к моменту появления зеленых листьев, очевидно, обусловлено уменьшением к этому времени интенсивности J-фактора.
Отметим, что чувствительность к J-фактору насыщенных водой тканей камбия и флоэмы будет возрастать в период весеннего пробуждения деревьев при низких положительных температурах по ночам (1 – 5оС), поскольку при этих условиях в воде эффективно образуются динамичные хиральные кластеры, играющие важную роль в механизме активации биохимических реакций в растениях и семенах (стратификация) [9]. Кроме того, по ночам усиливается действие J-фактора неэлектромагнитной природы [4]. Сезонные вариации интенсивности J-фактора могут отвечать не только за пробуждение семян и растений от состояния зимнего покоя, но и за инициацию метаболических реакций, отвечающих за цветение и половое размножение растений. По-видимому, именно величиной ночного вклада J-фактора в физиологию растений определяется их фотопериодизм и разделение на «растения длинной и короткой ночи» [5].
На этапе пробуждения растения от зимнего покоя и в процессе активизации тканей камбия J-фактор может индуцировать спиральную анизотропию (градиенты) в радиальной диффузии живицы и в вертикальном потоке сахарозы, несущим фитогормоны по флоэме. Очевидно, что в условиях такой анизотропии кинетика метаболизма и пространственное распределение фитогормонов в меристеме камбия приобретут винтовую симметрию, что и обусловит формирование косослоя в структуре новых слоев клетчатки.
Роль живицы в метаболизме активных тканей камбия до сих пор не установлена [7]. Учитывая упрощенность структуры клеток флоэмы у хвойных пород по сравнению с лиственными [7], можно полагать, что за спиротропизм хвойных деревьев ответственна главным образом живица. В силу гидрофобности основных компонентов живицы наиболее вероятно ее участие именно в энергетике метаболизма меристемы. Этому могут способствовать оптически активные смоляные кислоты (Рис 4), содержание которых в живице достигает ~66 % масс. Благодаря смолам и скипидару живица может сенсибилизировать жидкую среду камбия к хиральной составляющей внешней биогенной энергии, способствуя тем самым активации хиральных центров молекул фитогормонов и других метаболитов [2].
Можно также
предположить, что участие живицы в процессе биогенизации внешних хиральных
квантов энергии хвойными (голосеменными) деревьями могло сыграть ключевую роль в
ранние геологические эпохи, характеризуемые повышенной интенсивностью потока
солнечного нейтрино. На этих этапах генезиса растений в них развился механизм
аккумуляции квантов нейтринной энергии, максимум эффективности которого
достигался в ночное время при активном участии земной коры [4]. Данное
предположение согласуется с тем фактом, что самое высокое давление живицы в древесине
наблюдают на рассвете [7].
С выходом энергетики Солнца на
стационарный режим в спектре его излучения стала превалировать электромагнитная
энергия, что и обусловило зарождение наряду с хвойными деревьями
покрытосеменных или цветковых растений. Усложнение морфологии древесины и
водная основа ее жидкой среды, очевидно, сопровождалось перестройкой механизма
аккумуляции нейтринной энергии и снижением ее роли в физиологии покрытосеменных.
У хвойных же деревьев механизм аккумуляции нейтринной энергии с участием
живицы, очевидно, действует до сих пор, поэтому в местах с повышенной ее
плотностью она может вносить существенный вклад в спиротропизм.
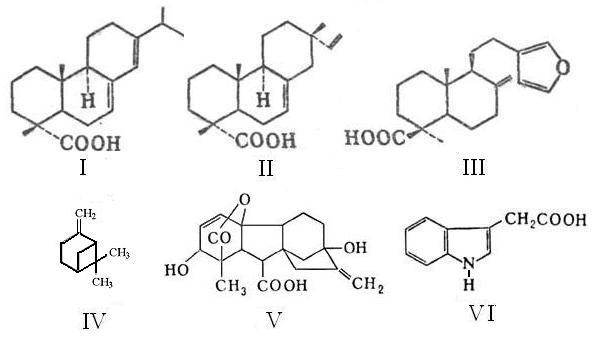
Рис 4. Структуры соединений составляющих основу живицы хвойных
деревьев (I – IV) и фитогормонов (V, VI). Смоляные кислоты: I – Абиетиновая
(–115о); II – изопимаровая (0о); III – ламбертиановая (+49о) (только
в сосне кедровой); IV – бета-пинен
(основа скипидара). В скобках – знак и удельный угол вращения. V –
гиббереллин; VI – ауксин.
Действие J-фактора заметно не только на уровне закрученности всего ствола в целом. Порождаемая его действием винтовая симметрия в динамике метаболитов неизбежно проявляется и на клеточном уровне морфологии древесины хвойных пород, однородной по составу и состоящей в основном из трахеид. Трахеиды отвечают за опорные и водопроводящие функции ксилемы. Пористые оболочки трахеид образуются из биополимера лигнина, третичная структура которого имеет вид колец или спиралей [5, 7]. Формирование такой метрики лигнина также может быть обусловлено действием J-фактора на этапе самоорганизации третичной структуры лигнина. При этом у некоторых хвойных деревьев образуется косослой ксилемы, и вода по ней начинает передвигается по спирали.
Спиротропизм изредка наблюдается в северном полушарии и у некоторых лиственных пород деревьев. Например, у дуба болотного ток воды проходит по спирали, а у каштана закрученность ствола проявляется даже на уровне коры [1]. Очевидно, эти случаи обусловлены аномально высокой интенсивностью J-фактора в местах произрастания таких деревьев. Винтовая симметрия метаболизма при этом может быть производной чувствительности к J-фактору, как транспорта фитогормонов по флоэме, так и радиальной диффузии пасоки по паренхимным клеткам, лучевые образования которых соединяют сердцевину с корой.
3.3. ФИЗИЧЕСКАЯ ПРИРОДА J-ФАКТОРА
С учетом известных и полученных данных можно полагать, что J-фактор, ответственный за спиротропизм деревьев генетически связан с ночной энергетикой, воздействующей на дерево через его корневую систему. Известно [5], что общая поверхность корней в 130 раз больше поверхности надземных частей того же самого растения, а их всасывающая площадь в 22 раза больше общей транспирирующей поверхности листьев. Такое отношение площадей надземной и подземной частей растения, очевидно, обусловлено различием интенсивностей обмена дерева энергией и веществом с атмосферой и почвой. В листьях в процессе фотосинтеза импульс фотона солнечного света, преобразуется в энергию химических связей, при этом СО2 дает углерод, а Н2О – водород.
Возникновение и развитие живых систем на начальных этапах эволюции осуществлялось в рамках «основного принципа действия» (ОПД) [10], требующего, чтобы на каждом шаге эволюции сохранялся баланс между действиями внешних поступательной и вращательной энергии, конкретные формы которых видоизменялись по мере возрастания сложности устройства живых систем [2, 10]. Для растений как одной из ранних форм живых систем форма проявления ОПД законсервировалась на достаточно примитивном уровне – в основе их метаболизма лежит сочетание двух вещественно-энергетических пар: (СО2 + импульс фотона) и (Н2О + момент импульса J-фактора), поступающих в дерево через крону и корни, соответственно.
Таким образом, потоку солнечных фотонов должен соответствовать исходящий из земли поток дискретных энергоформ (ЭФ), суммирующий в себе кванты полей различной природы (гравитационной, магнитной, нейтринной) [10]. Из данных квантов образуются квазинепрерывные соленоидальные трубки, подобные вихревым трубкам Фарадея, моделирующим силовые линии магнитного поля. Винтовое движение эфира по этим трубкам можно представить как перенос импульса и момента импульса. Базовая геометрия трубок ЭФ отвечает изотропной метрике гравитационного поля, которая модулируется метриками ЭФ магнитного поля и квантов нейтринной энергии [10]. Направление переноса импульса и момента импульса задается градиентом плотности вещества и для описания динамики потоков эфира применимы модели гидро- и электродинамики (уравнение Бернулли и явление самоиндукции) [10]. Очевидно, такая метрика потоков эфира, исходящих из недр земли, обусловила цилиндрическую форму элементов деревьев (корней, ствола, ветвей).
Условие неразрывности потоков ЭФ эфира в различных частях дерева можно выразить соотношением:
Niρi r2i = const,
где Ni – число волосков, корней, ветвей, стеблей листьев; ρi и r2i – плотности потоков эфира и радиусы соответствующих частей дерева; для ствола Ni = 1.
Очевидно, что некоторые из энергетических составляющих исходящего из земли потока эфира могут играть роль J-фактора. Прежде всего, отметим, что за вращательную компоненту энергетики метаболизма дерева и за природу J-фактора не может отвечать тепловая энергия почвы, которая представляет собой равновесное и изотропное излучение черного тела имеющего температуру почвы. Об этом же свидетельствует отрицательный результат в опыте по выявлению возможности хирализации теплового излучения оптически активной средой.
Маловероятно также, чтобы J-фактор, ответственный за спиротропизм, имел магнитную природу. В принципе, на двигающиеся по флоэме или ксилеме заряженные метаболиты будет действовать сила Лоренца, играющая роль центростремительной силы:
evzHR = mvz2/R или eHR = mvz/R, (1)
vz – вертикальная составляющая скорости движения метаболита, m – его масса (~10–25 кг), е – единичный заряд (~10–19 Кл), HR – горизонтальная составляющая магнитного поля земли (~10–5 Тл), направленная по радиусу (R ~ 0,1 м) ствола дерева. Для этих значений из (1) получается величина vz ~ 1м/c, что на три-четыре порядка выше скоростей перемещения метаболитов по ксилеме и флоэме деревьев (см. П. 3.2). Причем, отсутствие спиротропизма для лиственных деревьев, тогда как для них vz на порядок выше, чем для хвойных также указывает на отсутствие заметного эффекта силы Лоренца на метаболизм деревьев. Для ощутимого влияния аномального магнитного поля на магнитные моменты метаболитов его значение должно достигать величин на четыре порядка превышающих значение магнитного поля земли.
Механизмы геотропизма различаются для корневой и надземной частей дерева. Опыты по выращиванию растений на космической станции в условиях невесомости показали, что замена силы тяжести на равнозначную ей силу инерции восстанавливает вегетативный цикл растения, но при этом функция полового размножения претерпевает специфические изменения [11]. Меняется химический состав пахучих веществ цветущей розы, а семена пшеницы, вызревшие в космосе имеют сдвиги на уровне генома. Учитывая эти результаты и отсутствие адекватной теории гравитационных взаимодействий нельзя полностью исключить возможность их вклада в формировании вращательной составляющей энергетики метаболизма и J-фактора.
В работе [2] на роль внешнего фактора инициирующего формирование ресурса функциональной асимметрии у человека в процессе ночного сна были предложены вихревые возмущения стационарного потока солнечного нейтрино, интенсивность которого составляет порядка 3% от интенсивности электромагнитного излучения Солнца. Очевидно, такое соотношение двух биогенных форм солнечного излучения и обусловило превалирующее развитие у растений корневой системы для соблюдения баланса вкладов в метаболизм растений нейтринной (ночной) и световой (дневной) энергиями. Источником возмущений стационарного потока нейтрино, по-видимому, являются вихревые магнитные трубки в фотосфере, которые проявляются на поверхности Солнца пятнами. Вихревые возмущения потока нейтрино пронизывают и фокусируется Землей на ее ночной поверхности и приводят к различным аномальным явлениям в биосфере. У людей эти импульсные возмущения могут инициировать творческую деятельность и скачки в духовной эволюции общества [3], а у хвойных деревьев локальные вихри нейтринного потока вполне могут играть роль J-фактора, ответственного за спиротропизм.
Как было отмечено в [2, 3], концентрации и биогенизации вихревого потока нейтрино способствует керамзит. В почве такую функцию могут выполнять силикаты, которые составляют от 30 до 80% от общей массы почвы. Они являются энергетической матрицей почвы и оказывают влияние практически на все ее свойства [12]. Большой процент в силикатах составляют соединения двуокиси кремния в основном в виде микрокристаллов оптически активного кварца. Переносу квантов нейтринной энергии от кристаллов кварца на хиральные структуры корневых волосков будет способствовать почвенная вода и сахар, содержащийся в слизи, выделяемой волосками и кончиками корней.
Согласуются с предложенным механизмом обогащения корневой системы растения нейтринной энергией автоопыт по лечению радикулита и синовита голеностопного и локтевого суставов с помощью песочных ванн на побережье Красного моря. Основной причиной синовита, очевидно, является снижение упругости синовиальной жидкости из-за низкой плотности конденсата нейтринной энергии на гиалуроновой кислоте, отвечающей за динамические свойства синовии. При погружении тела в песок (Т ~ 40 – 50оС) прилегающий к телу слой песка смачивается потом, который состоит на 89-90% из воды и широкого набора метаболитов, в том числе и хиральных. Увлажненный слой песка играет роль концентратора для квантов нейтринной энергии, переходящих от него на хиральные метаболиты в синовии и спинномозговой жидкости. Четырех процедур длительностью ~15 мин (утром и вечером) оказалось достаточно для устранения боли в поясничном отделе позвоночника и суставах.
Снижение плотности стационарного потока солнечной нейтринной энергии может приводить к уменьшению корневого давления и как следствие этого к падению интенсивности водоснабжения дерева. Особенно чувствительным в этом смысле оказываются дубы с их кольцесосудистой системой водоснабжения, с высотой сосудов у зрелых дубов достигающей трех метров. Эффективная работа такой системы требует высоких значений корневого давления, которое в свою очередь определяется плотностью стационарного потока нейтринной энергии. С его снижением в ХХ столетии [4] можно связать глобальное явление усыхания зрелых дубов в северном полушарии, объяснения которому до сих пор нет [13].
4. ЗАКЛЮЧЕНИЕ
Известные и экспериментальные данные свидетельствуют о наличии специфических реакций на действие внешних физических факторов у хвойных и лиственных пород деревьев. Для первых характерен аномальный спиротропизм (закручивание структуры древесины вокруг оси дерева), а для вторых – аномальный электротропизм, конкурирующий с геотропизмом при определенных условиях. Данные различия в механизмах адаптации деревьев к внешним физическим факторам обусловлены разницей в физико-химических свойствах жидкой среды и морфологии лиственных и хвойных пород. Солнечное нейтрино в случае его биогенности вполне может играть роль внешнего хирального фактора ответственного за аномальные явления спиротропизма.
ЛИТЕРАТУРА
1. Меннинджер Э. Причудливые деревья. – М.: Мир, 1970, - 371 с.
dendrology.ru/books/item/f00/s00/z0000004/st019.shtml
2. Холманский А.С. Дихотомия правого и левого в живых системах //
www.j-symmetry.com/Holmansky_03_08.htm; Зависимость функциональной асимметрии мозга от внешних условий // www.j-asymmetry.com/Kholmansky_1_09.htm
3. Холманский А.С. Физические факторы духовной эволюции // www.smolensk.ru/user/sgma/MMORPH/N.../holmansky-2.doc
4. Холманский А.С. Биогенность нейтрино // Сознание и физическая реальность, 2004, №4, С.35; Кинетический фактор фотосинтеза // http://kubstu.ru/fh/fams/kinetika.doc.
5. Полевой В.В. Физиология растений, - М.: Высшая школа, 1989, - 464 с.
6. Гордеев А.М., Шешнев В.Б. Электричество в жизни растений, М.: Наука, 1991, 160 с
7. Физиология древесных растений // www.bonsai.ru/dendro/phcontent.html
8. Артамонов В.И. Занимательная физиология растений, -М.: Агропромиздат, 1991. -336 с.
9. Холманский А.С. Особенности термодинамических свойств воды и биоэнергетика // Доклады РАСХН. №2. 2006. С. 63; http://www.sciteclibrary.ru/rus/catalog/pages/7897.html
10. Холманский А.С. Самоиндукция эфира // SciTecLibrary - Cтатьи и Публикации; Метрика приземного эфира // http://www.sciteclibrary.ru/rus/catalog/pages/7876.html;
11. Нечитайло Г. С. Форум журнала Российский космос. №1. – 2006
12. Поздняков А.И. Полевая электрофизика почв. -М.: МАИК, 2001, 187 с.
13. Причины деградации дубрав в мире, сайт www.treeland.ru
ADAPTATION OF TREES TO ABNORMAL PHYSICAL
FACTORS
Holmansky A. S.
Presence of specific reactions to action of external physical factors at
coniferous and deciduous breeds of trees is established. For the first it is
characteristic abnormal spirotropism (a twisting of structure of wood around of
an axis of a tree), and for the second – abnormal electrotropism, competing
with geotropism under certain conditions. The given distinctions in mechanisms
of adaptation of trees to external physical factors have connected with
differences electrophysical and chiralyc properties of the liquid environment
and morphology of deciduous and coniferous breeds. Using a hypothesis about
biogenic solar neutrino, have offered the possible mechanism of its action on
trees as external chiral the factor responsible for the phenomenon spirotropism.
Keywords:
trees,
electrotropism, spirotropism, abnormal fields, neutrino.
ВНИИ электрификации сельского хозяйства (РАСХН), Москва
Поступила в редакцию 9.08.2009.