Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 9. -
Вып. 4. - 2010. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-28-html/TITL-28.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-28-html/cont.htm
УДК 576.1
ХИРАЛЬНОСТЬ
И КВАНТОВЫЕ ЭФФЕКТЫ КАК ФАКТОРЫ МОРФОГЕНЕЗА
Ó 2010 г. Холманский А. С.
В обзоре проведен анализ
данных по хиральной дихотомичности как фактору, направляющему морфогенез по
стреле времени – от простого к сложному. Представлены аксиоматика и правила
применения универсального математического языка, позволяющего единообразно
описывать закономерности изменения дискретных форм материи и квантовых состояний
биосистем на всех уровнях их организации от субэлементарного до социально-духовного.
Проанализирована степень хиральности анатомических и функциональных элементов
физиологии человека, образующих квантовые когерентные ансамбли, чувствительные
к внешним физическим факторам хиральности, как электромагнитной, так и
нейтринной природы. Обсуждена роль квантовых свойств воды в биоэнергетике и
процессах самоорганизации когерентных ансамблей. Руководствуясь логикой антропного принципа, вектор эволюции
отождествили с градиентом хиральности и
связали с ним степень хиральной чистоты мозга, с которой соотнесли
функциональную асимметрия мозга и эффективность эвристического мышления.
Ключевые слова: биосистема, морфогенез,
энергоформа, квантовый, асимметрия, ядерный спин, вода, мозг.
ВВЕДЕНИЕ
Вектор эволюции реализуется как закономерный процесс
изменения формы, содержания и поведения биологической системы (биосистемы) под
действием меняющихся физических факторов различной природы. Понятие биосистемы
относится к отдельным живым организмам и ко всей биосфере. Антропный принцип
(АП) свидетельствует, что программа изменения гелиогеофизики имеет своей целью
зарождение жизни на Земле и формирование двудольного мозга человека с
последующим развитием его хиральной
дихотомии вплоть до кооперирования мыслительных способностей отдельных людей в
единую ментальную систему (метамозг).
Поскольку любое действие квантовано [1], а всякая
система – дискретна, то и эволюция, в принципе, является скачкообразным или квантовым
процессом. Закономерность эволюции (номогенез
[2]) есть следствие подчинения внутри- и межсистемных связей универсальным
физическим законам. Соответственно, физическая природа внешних факторов будет
определяться природой тех связей, которые наиболее чувствительны к изменению
внешних условий.
Квантовый морфогенез, как и парадигма АП [3] опирается
на достоверные данные атомно-молекулярной физики. Стандартная теория
элементарных частиц в силу своей ахиральности и абсурдности [4] в принципе не
годится для решения фундаментальных проблем номогенеза. Поэтому изучение
механизма участия в морфогенезе факторов внешней среды до сих пор остается на
этапе накопления достоверных эмпирических данных и не имеет адекватной
теоретической базы. Понятно, что подразделение факторов морфогенеза на
внутренние и внешние условно в силу многоуровневой «открытости» реальной биосистемы
и отсутствия понимания физической природы слабых
взаимодействий в биосистемах [4, 5]. Решение этого вопроса возможно при
адекватной формализации синергизма внутренних и внешних факторов эволюции с
помощью универсальных физических понятий, позволяющих единообразно выражать
механизмы действия факторов на всех уровнях организации биосистем.
Универсальный язык, позволяющий адекватно описать
квантовый морфогенез построили с помощью следующих фундаментальных определений
[1]:
- аксиомы дух
есть сущность материи;
- понятия энергоформа
(ЭФ);
- логики основного
принципа действия (ОПД).
Согласованность аксиоматики и логики данного языка с
законами диалектики (единства и борьбы противоположностей, подобия) есть
гарантия его внутренней непротиворечивости. Аксиома позволяет экстраполировать
семантику языка на духовную сферу, применяя закон подобия в виде
духовно-физического изоморфизма [6]. Данное обстоятельство дает возможность
представить процесс формирования метамозга как духовный этап морфогенеза.
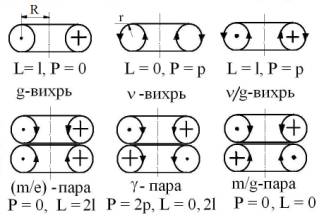
Рис 1. Схемы базовых ЭФ. Изначальные вихри эфира
(духа): g- вращающийся, L – момент импульса; ν- самодвижущийся, Р –
импульс; ν/g- винтовой (правый и левый)
и их стабильные пары: m/e-покоящаяся (заряд, масса
покоя); γ- движущаяся (эквивалентная масса фотона); m/g-
покоящаяся («скрытая материя»).
Язык квантового морфогенеза имеет свою «азбуку» и
«грамматику» [1]. «Знаками азбуки» являются:
- набор базовых ЭФ (Рис 1);
- самосогласованные численные значения мировых
констант (постоянная Планка – h = 6,67 10–34
Дж с, число Авогадро – N = 6,02 1023
1/моль, скорость света – С = 108 м/с).
«Грамматика» языка включает:
- правила сочетания базовых ЭФ в более сложные ЭФ с
соблюдением ОПД и сборки из этих ЭФ квантов полей различной природы, а также
дискретных элементов структур частиц и ядер – оболочек и орбиталей;
- принципы определенности, выражающие квант действия
через произведение трех пар взаимосвязанных физических величин (обозначения
поясняются на рисунке): энергии и времени ее действия (E t), импульса и
шага перемещения – λ = 2πr (Р
λ), момента импульса и оборота на угол 2π (L 2π);
- формализм фрактально-резонансных изоэнергетических обратимых
трансформаций или флуктуаций ЭФ эфира:
![]() (1)
(1)
В (1) вихрь с характерным радиусом потоков эфира в его
атмосфере rn преобразуется в k вихрей с радиусом krn (раскрутка) и обратно (конденсация). Вихри, образуя
спираль, могут порождать силовую трубку соленоидального поля или замыкаться в
тор. Из таких трубок и торов формируются силовые линии электромагнитного (ЭМ)
поля. Отрыв n « k звеньев от
спирали можно представить как излучение кванта эфира, переносящего импульс и
момент импульса. Кинетика флуктуаций базовых ЭФ может быть ограничена
константой скорости СN1/2, а действие квантов эфира ЭМ-природы константой С.
Базовые ЭФ позволяют смоделировать изначальную форму
материи, заполняющую пространство эквивалентной массой или потенциальной
энергией. В зависимости от ориентации векторов P и L ЭФ могут быть
правыми (вектора параллельны) и левыми (вектора антипараллельны). Хиральность
комбинированных ЭФ определяет хиральность квантов физических полей,
элементарных частиц и ядер, внутренняя структура которых автоматически
удовлетворяет ОПД.
С помощью (1) можно
формализовать стационарные флуктуации ЭФ реликтового эфира, которые проявляются
микроволновым излучением космического газа, находящегося при температуре 2,71К.
Эта величина не случайно равна
основанию натурального логарифма – е [7].
Температура межзвездной
среды является совокупной мерой количества движения ~N ЭФ, занимающих
определенный объем пространства. Применительно к реликтовому фотону параметр Т будет равнозначен его импульсу (Е =
РС ~ kT) или кинетической энергии, которая будет равна суммарной Еn-энергии порядка N ЭФ
ЭМ-природы, конденсирующих на молекуле межзвездного газа по (1). Энергетический
спектр реликтовых ЭФ при Т ~ 0 отвечает формуле Бозе распределения:
n
= 1/[ехр(Еn/kT) - 1 ] .
Преобразовав это выражение к тождественному виду:
ехр(Еn/kT) = 1 + 1/n
и
учтя:
e = lim (1 +
1/n)n = 2, 71, при
n ® ¥ ,
для n порядка N получим соотношение:
РС =
NЕn ~ kT. (2)
Длину волны реликтового фотона можно оценить по
формуле Вина:
λ = b/T ~ 1, 1 мм (3)
где
b – постоянная Вина, равная 2,9 10–3 м К, а
Т = 2,7К. Движение фотона обеспечивается импульсом (Р), которому соответствует
самодвижущийся вихрь эфира с характерным радиусом rn =
λ/2π = 0,175 мм и импульсом
Р = h/λ = ħ/rn. (4)
Радиус реликтовых ЭФ, конденсирующих в структуру
фотона и образующих «корону Гамова» [7], равен Nrn ~ 1020
м, то есть одного порядка с радиусом Галактики. Отметим, что по такой же схеме
происходит k-конденсация ЭФ (k ≤ N) в
живых системах в процессе их самоорганизации. При этом квант ЭМ-энергии может
инициировать какой-нибудь физико-химический акт (процесс канализирования
энергии) или излучиться системой во внешнюю среду. В последнем случае энтропия
системы понизится на величину q/T, где q = РС – квант
тепловой энергии системы [4].
Флуктуации реликтового эфира универсальны и
повсеместны, об этом свидетельствует Лэмбовский сдвиг в энергии электрона атома
водорода. Можно предположить, что ритм данных флуктуаций инициирует конденсацию
соответствующих ЭФ, например, в однородных системах нервных клеток, входящих в
пейсмейкеры (пучок Гиса, ритмоводители мозга). В общем случае масштабом и уровнем
структурной однородности биосистемы задается радиус и тип слабых
взаимодействий, формирующих когерентную квантовую систему, в которой работает
схема (1).
Притяжение или отталкивание
между частицами, имеющими заряд и момент импульса, в принципе, можно объяснить,
экстраполируя кинематику и динамику газовых или жидкостных вихрей на
вихреподобные ЭФ эфира и соленоидальные поля, имеющие источники и стоки.
Отметим, что аналогично ЭМ-силам действуют ядерные силы, за них ответственны
потоки хиральных ЭФ эфира, генерируемые нуклонами. Взаимодействия данных ЭФ
представляют как «переносимое глюонами цветовое взаимодействие между составляющими
элементами нуклонов (кварками)» [8]. Глюоны здесь соответствуют ЭФ поля ядерных
сил, цветность – хиральности потоков ЭФ, а кварки – оболочкам и орбиталям нуклонов
[1].
Язык ЭФ позволил рассчитать
модели структур пяти элементарных частиц (нейтрон, протон, электрон, нейтрино,
фотон) и их возбужденных состояний [9], которые по недоразумению до сих пор [4]
относят к нестабильным элементарным частицам. Рассчитали также структуры легких
ядер и Солнца [7]. Все перечисленные структуры, в принципе, изоморфны структуре
протона (Рис 2) и различаются числом орбиталей и направлением спина элементов,
определяющим спин или хиральность. К примеру, сумма спинов соответствующих
элементов дает положительное значение спина у протона и отрицательное у
электрона и нейтрона.
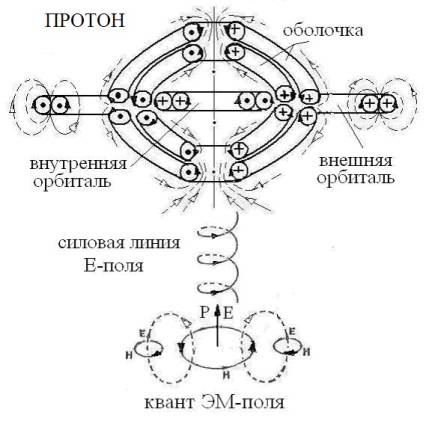
Рис 2. Схема структуры протона и акта поглощения протоном
правого кванта ЭМ-поля (ЭМ-ЭФ). Сплошные линии относятся к замкнутым потокам g-вихрей
(магнитное поле – Н); пунктирные - ν-вихри (электрическое поле – Е). Квант ЭМ-поля моделирует принцип связывания
ν- и g-вихрей в сложных ЭФ.
Ядра нуклоноподобны и собираются из многослойных
концентрических оболочек и орбиталей. Гибридизированной геометрией ЭМ-поля ядра
определяются конфигурации атомных электронных орбиталей. Таким образом,
квантование внутренней структуры ядер предшествует закономерностям изменения
физико-химических свойств элементов в
таблице Менделеева. Формирование и изменение геометрии молекулярных электронных
орбиталей также подчиняется законам квантовой механики. Поэтому поведение
биосистем квантуется как на уровне метаболизма, так и на ментально-социальном
уровне.
1. АСИММЕТРИЯ МОРФОГЕНЕЗА
В силу внешних по отношению к миру причин правые
производные базовых ЭФ оказалась более устойчивы, что и обеспечило
доминирование их активности при формировании реликтового состояния эфира на начальном
этапе самоорганизации Вселенной [1, 9]. Это, очевидно, и обусловило анизотропию
фоновой динамики эфира, которая в дальнейшем стала играть роль универсального
фактора хиральности пространства (ФХП), ответственного за самоорганизацию Вселенной
из частиц, а не из античастиц (барионная асимметрия).
Роль ФХП сохранилась и после формирования
вещественного костяка Вселенной, гравитационные и ЭМ-поля которого
соответствующим образом деформировали метрику реликтового эфира. Можно
полагать, что ФХП, влияя на слабые взаимодействия в биосистемах, определяет
направление их развития по стреле времени, то есть от простого к сложному.
Адекватным параметром сложности организации биосистемы будет степень ее чувствительности
к ФХП. Соответственно, эволюцию по стреле времени будет характеризовать уровень
хиральной чистоты самого сложного вещественного образования Вселенной –
двудольного мозга человека.
Хиральная чистота мозга, определяя меру его
чувствительности к ФХП, проявляется,
прежде всего, как функциональная асимметрия мозга (ФАМ). ФАМ сочетает в себе
дифференциацию и синергизм функций правой и левой доли мозга и лежит в основе
механизма эвристического мышления. Усложнение морфо-физиологических проявлений
ФАМ в процессе эволюции равнозначно развитию умственных способностей человека
вплоть до формирования механизма их кооперирования в единую интеллектуальную
систему (метамозг).
Функционирование и единосвязность всех уровней
биосистем обеспечивают ЭМ-взаимодействия в широком диапазоне энергий. Его реализуют
фотоны (от ультрафиолетового до КВЧ-диапазона), фононы, магноны, а также ЭМ-ЭФ
и ЭФ нейтринной природы (Х-ЭФ) [5, 8]. Функциональные взаимодействия элементов
биосистемы, целевые и неравновесные по своей сути, осуществляются за счет
физико-химических механизмов канализования тепловой энергии (kT) когерентными ансамблями однородных элементов. Механизмы
канализования зависят от структуры и динамических характеристик элементов и
сплошных сред организма. Хиральные среды и метаболиты сообщают кинетике процесса канализования чувствительность к ФХП. По сути,
хиральный фактор кинетики неравновесных процессов в когерентных подсистемах
суммирует в себе асимметрию динамичных и электрохимических свойств метаболитов,
тканевых структур, органов и жидких сред биосистемы.
Вода как матрица и активный метаболит играет ключевую
роль в механизмах самоорганизации биосистем [10]. Это связано, прежде всего, с
ее аномальными физическими свойствами, имеющими квантовую природу [11]. В
настоящей работе, используя язык и понятия квантового морфогенеза,
проанализировали возможные механизмы реализации его хиральной доминанты в
физиологии человека.
2. ГРАНИЦЫ БИОЭНЕРГЕТИКИ
Метаболизм, рост и развитие организма человека
реализуется, прежде всего, за счет действия квантов ЭМ-энергии при
посредничестве ЭМ-ЭФ, генерируемых взаимодействующими и подвижными частицами.
Реальные заряженные частицы (ионы, ион-радикалы, полярные молекулы) помимо
заряда или дипольного момента имеют момент импульса (спиновый или орбитальный).
Броуновские трансляционные, колебательно-вращательные и конфирмационные
хаотические движения частиц или фрагментов биомолекул предполагают наличие
стационарного обмена квантами тепловой энергии (Т-фотоны), больцмановское
распределение которых по частотам отвечает энергетическому спектру всех движений.
Стационарная плотность и энергия Т-фотонов в среде
хаотически распределенных частиц однородной системы отвечает значению ее
локальной температуры. Длину волны Т-фотона можно оценить по закону Вина (2):
λ = b/T = 9,4 мкм, а rn = 1,5 мкм
при Т = 310К (средняя температура организма – 36,6оС). Параметры
ЭФ, обеспечивающей движение Т-фотона, с учетом (1), позволяют рассматривать ЭФ
в качестве движителей метаболитов в нервных волокнах и жидких средах, включая
кровеносные капилляры и поры клеточных мембран.
Импульс и момент импульса Т-фотонов и ЭФ с
учетом возможности их трансформации по схеме (1) и механизма канализования
обеспечивают работу всех динамических подсистем организма (гуморальной, нервной,
мышечной). Первичным физико-химическим актом является перенос электрона или
протона. Энергетический спектр фотонов как экзо-, так и эндогенных определяется
видами движений и характерными метрическими параметрами когерентных электронных
ансамблей. Сплошность жидкой среды кровеносной и нервной систем и наличие в ней
электролитов позволяют считать их односвязными электрофизическими системами с
метрическими характеристиками от долей микрона до метра.
Нижняя граница спектра Т-фотонов отвечает энергии крутильных
колебаний отдельной молекулы как целого в поле окружающей среды [12]. Например,
для воды наблюдались колебания в области 15-85 см–1 и 200-600 см–1
[12], а энергия возбуждения ее кластерных структур составляет ~10 Дж/моль.
Верхняя граница экзогенного Т-фотона соответствует тепловой энергии или энергии
активации трансляционной подвижности свободной молекулы воды ~2,5 кДж/моль
[10]. Этой энергии вполне достаточно для обеспечения стационарной биоэнергетики
здорового организма.
Можно также полагать, что при канализовании тепловой
энергии в процессе самоорганизации когерентных ансамблей могут идти побочные
физико-химические акты, приводящие к возбуждению электронных состояний
метаболитов, с последующим излучением фотонов видимого и УФ-диапазона. Примерами
таких процессов служит трибо- и хемилюминесценция (светлячки), а также
митогенетическое излучение Гурвича [13]. В основе этих процессов лежат реакции
рекомбинации ион-радикалов. Их источником может быть ионизирующее излучение
радиоактивных экзо- и эндогенных элементов, а также воздействие на
живые ткани раздражителей – наркотических средств,
механических и электрических импульсов,
резкого охлаждения. Как правило,
такие реакции инициируют появление тех или иных патологий.
Диапазон энергии эндогенных фотонов, воздействие
которых на организм не приводит к патологическим реакциям, несколько шире
диапазона энергии экзогенных Т-фотонов, поскольку включает в себя фотоны
возбуждающие зрительную систему (до 350 нм, ~400 кДж/моль), а также инфразвук и
радиоволны (до ~1 м или ~0,1 Дж/моль).
Основным источником экзогенных Т-фотонов является
химическая реакция ферментативного окисления глюкозы, тепловой эффект которой
аккумулируется затем на электронной системе молекулы АТФ в виде связанных
Т-фотонов [10]. Механизмы высвобождения и действия связанных фотонов в
метаболизме до сих пор не установлены [14]. Очевидно, что излучательный и
безизлучательный механизмы переноса ЭМ-энергии в значительной степени
определяются стереохимией метаболитов и молекулярно-клеточных структур, а также
динамикой водородных связей в сплошных средах. При этом механизмы миграции
энергии зависят, прежде всего, от степени когерентности ансамблей или от интенсивности
корреляционных взаимодействий в них на уровне ЭФ [14].
3. ХИРАЛЬНАЯ ГИСТОЛОГИЯ
В основе структурной и функциональной асимметрии
биосистем лежит хиральность спирального движения частиц и квантов энергии.
Кроме того, биомеханику на макро- и микроуровне лимитирует реология жидких или иных
однородных, сплошных сред организма, которые из-за обязательного присутствия в
них оптически активных веществ или молекулярно клеточных структур можно считать
хиральными средами. Типичными представителями оптически активных метаболитов и
структур будут белки и сахара в крови, гиалуроновая и молочная кислота в
стекловидном теле глаза и синовии, в соединительных и мышечных тканях;
коллагеновые волокна в дерме кожи и в костных тканях. Рассмотрим известные
данные по асимметричным молекулярно-клеточным структурам, играющим важную роль
в биоэнергетике и сенсорике организма.
Примером действия хирального фактора на молекулярном
уровне может служить механизм ферментативного синтеза АТФ в матриксе
митохондрии. Активация фермента АТФ-синтазы достигается за счет энергии
протонного градиента на мембране [15]. Поток протонов инициирует вращение
одного блока фермента (F0), относительно другого (F1) и при
этом осуществляется присоединение к аденозиндифосфату (АДФ)
фосфата.
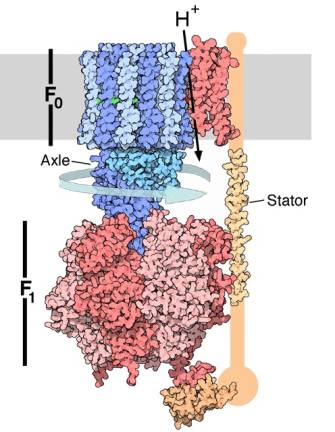
Рис 3. Структура АТФ-синтазы. Протонный канал F0
и вращающаяся часть показаны синим, компонент F1 — красным, мембрана
— серым [15].
Механизм поворота и
взаимосвязь между знаком градиента протонов и направлением вращения блока фермента
АТФ-синтазы до конца не изучены. Можно предположить, что протонный ток
поляризует в блоке F0 α-спирали белков, они смещаются вдоль его оси и
при этом спиральность белков вынуждает блок поворачиваться, как в червячной
передаче.
Хиральность сенсорно-информационной
сферы организма может быть обусловлена
спиральной структурой миелиновых оболочек нервов ЦНС. До сих пор открыт вопрос
о знаке этих спиралей и его влиянии на механизм распространения потенциала
действия. В работе [16] предположили, что спиральные насечки в миелиновой
оболочке нерва (Рис 4) участвуют в сальтаторном механизме проводимости нервного
возбуждения.
Электрофизика
механизма участия насечек в ретрансляции ЭМ-сигнала по сегментам нерва
родственна электрофизике потовых каналов, которые сообщают поверхности кожи
свойство антенны, способной резонансно поглощать и излучать фотоны
КВЧ-диапазона [17]. Спиральные насечки в миелиновых оболочках изоморфны потовым
каналам и наполнены слабым электролитом, поэтому они, аналогично спиралям
потовых каналов (Рис 5), могут взаимодействовать с вихревыми экзогенными
ЭМ-полями, генерируемыми мембранными и цитоплазматическими токами в перехватах
Ранвье [16]. Резонансные частоты насечек, очевидно, будут определяться их
метрическими и диэлектрическими характеристиками.
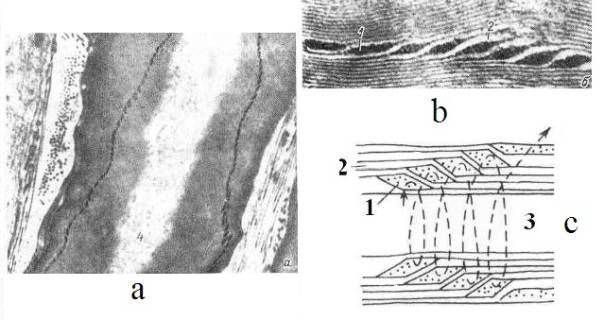
Рис 4. Ультраструктура миелиновой мембраны нерва с
насечкой (a, b) и схема насечки в миелиновой оболочке аксона (с) [16].
Установлено [18], что
у большинства людей до ~90% спиралей потовых каналов являются правыми (Рис 5).
Данная асимметрия может быть следствием хиральности среды эпидермы, содержащей
оптически активные вещества и структуры, и чувствительной поэтому к ХФП. Доминирование правых спиралей в морфологии
потовых каналов служит косвенным подтверждением гипотезы о важности согласования
знака спирали миелиновой оболочки нерва с направлением распространения
потенциала действия. К примеру, электрические импульсы, отвечающие болевым
раздражениям, могут двигаться по нервному окончанию с правой закруткой миелина
благодаря импульсу левоспиральных ЭМ-вихрей [16]. Кроме того, инверсные знаки
спиральности миелиновых оболочек черепно-мозговых и ассоциативных нервов,
локализованных в правом и левом полушариях, могут обеспечивать дифференциацию
их функций в рамках ФАМ и, прежде всего, на уровне психофизиологии. Рацемизация
или патологическая инверсия знаков спиральности у сигнальных и ассоциативных
связей внутри и между полушариями может проявиться в таких дисфункциях
психофизиологии и моторики, как леворукость и гомосексуализм.
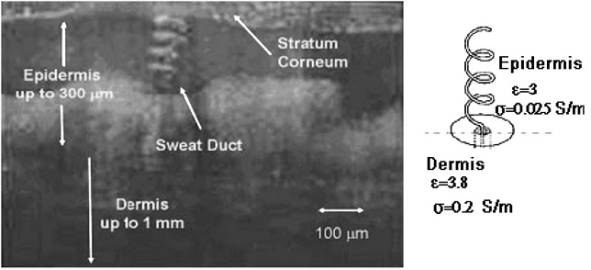
Рис 5. Трехмерная микрофотография и схематическое
изображение спирали потового канала [17]
Патологические инверсии хиральности миелиновых
оболочек черепных нервов могут закладываться на этапе зачатия или эмбриогенеза
под влиянием внутренних и внешних факторов. К первым относятся: химические
вещества (алкоголь, наркотики, лекарства) и стрессовые состояния. К внешним
факторам следует отнести, прежде всего, изменения геокосмических условий,
влияющие на знак ХФП и биоэнергетику организма [19].
4. КВАНТОВАЯ РЕОЛОГИЯ
Вода составляет основу крови, цитоплазмы и
межклеточной жидкости, ликвора и синовии. Во всех этих жидкостях в той или иной
концентрации присутствуют неорганические вещества, белки, сахара клеточные
образования, имеющие соответствующие гидратные оболочки. Кровь является
единственной подвижной тканью, которая течет только по сосудам. Ее следует
рассматривать как гетерогенную многокомпонентную систему корпускулярной
природы, содержащую форменные элементы (эритроциты, лейкоциты, тромбоциты),
которые находятся во взвешенном состоянии в коллоидном растворе электролитов,
белков и липидов. Кровоток представляет собой прохождение концентрированной
суспензии эластических дисков через трубки микрососудов, диаметр просвета
которых в их капиллярной части в ряде случаев намного меньше диаметра этих
дисков. В этих условиях на макроуровне могут проявляться квантовые свойства
молекул воды и веществ крови, поэтому капиллярную гемодинамику условно назовем квантовой реологией.
В принципе, квантовая реология лежит в основе всей
биоэнергетики организма. В настоящей работе рассмотрим хиральные
межмолекулярные взаимодействия на примере реакции ассоциирования сахаров и
проанализируем роль вращательной подвижности молекул воды в квантовой реологии.
На Рис 6 показаны зависимости удельного угла вращения ![]() декстрана и скипидара от температуры и даты
[19]. Снижение
декстрана и скипидара от температуры и даты
[19]. Снижение ![]() раствора декстрана при жаркой погоде в июле
августе в Подмосковье обусловлено с распадом молекулярных комплексов, имеющих
более высокое значение удельного вращения, по сравнению с олигомерами декстрана
[10]. Энергия активации реакции образования комплексов для различных сахаров
меняется в пределах 0,1 – 0,6 кДж/моль и изменение температуры на 10оС
уже дает ощутимый эффект [10].
раствора декстрана при жаркой погоде в июле
августе в Подмосковье обусловлено с распадом молекулярных комплексов, имеющих
более высокое значение удельного вращения, по сравнению с олигомерами декстрана
[10]. Энергия активации реакции образования комплексов для различных сахаров
меняется в пределах 0,1 – 0,6 кДж/моль и изменение температуры на 10оС
уже дает ощутимый эффект [10].
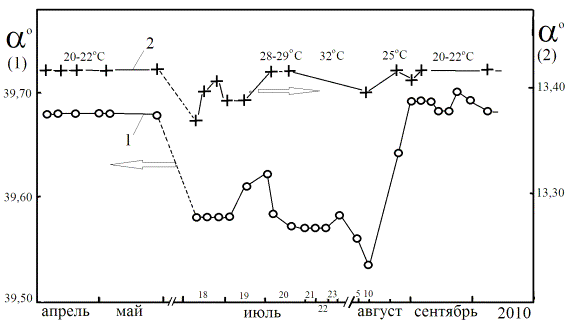
Рис 6. Зависимость угла вращения (α) оптически
активного раствора декстрана (кювета 200 мм) – 1) и скипидара (кювета 100 мм) –
2) от даты и температуры внешней среды [19].
Молекулы скипидара не образуют комплексов, поэтому его
угол вращения не реагирует на перепады температуры. Незначительные синхронные
изменения величины ![]() раствора декстрана и скипидара, например, 21
июля и 9 августа могут быть обусловлены изменением магнитной обстановки на
Земле вследствие повышения активности Солнца – в эти дни существенно возрастала
суммарная площадь солнечных пятен и возрастала интенсивность радиоизлучения на
длине волны 10,7 см (по данным ИЗМИРАН).
раствора декстрана и скипидара, например, 21
июля и 9 августа могут быть обусловлены изменением магнитной обстановки на
Земле вследствие повышения активности Солнца – в эти дни существенно возрастала
суммарная площадь солнечных пятен и возрастала интенсивность радиоизлучения на
длине волны 10,7 см (по данным ИЗМИРАН).
Экстраполируя
данные по зависимости оптической активности раствора декстрана от температуры и
электромагнитных возмущений на родственные хиральные среды организма можно
полагать, что межмолекулярные взаимодействия хиральных метаболитов в жидких
средах вносят свой вклад в чувствительность организма к ХФП.
Аномальные свойства воды, во многом определяющие
биофизику жидких систем организма, связаны с квантовыми эффектами водородной
связи, как в объеме воды, так и в гидратных оболочках метаболитов и на стенках
сосудов. В силу структурирования воды водородными связями
вращательно-ориентационные движения молекул воды, играющие важную роль в
процессах самоорганизации, имеют характер либраций (крутильных колебаний) и
существенно зависят от локальных электромагнитных полей. Последние могут играть
роль возмущений, снимающих симметрийные
запреты на возбуждение либраций Т-фотонами.
В общем случае вероятность возбуждения вращательного
состояния молекулы воды определяется интенсивностью взаимодействия ее
дипольного момента с соответствующим Т-фотоном, а энергия и момент импульса
пропорциональны ее моменту инерции. Свой вклад в правила отбора переходов и в
величину момента импульса вносит суммарный спин протонов молекулы, который
может быть равен 0 (пара-изомер) и 1 (орто-изомер). У свободной молекулу воды момент инерции имеет три значения (Рис
7), два из которых (J1, J3) сильно возрастают при закреплении молекулы на какой-либо заряженной поверхности
или молекуле [20].
Равновесное
орто/пара отношение для свободных молекул воды, например в воздухе, при комнатной температуре согласно квантовой
статистике равно 3:1. Часть параизомеров
Н2О находится на основном уровне и не вращается, в то время как
основной вращательный уровень
ортоизомеров смещен на 23,8
см-1 от нулевого
уровня (kT~ 0,28 кДж/моль, Т ~34
К), поэтому свободные ортоизомеры
вращаются всегда, пока заселен этот и выше лежащие уровни [21]. Поскольку орто/пара конверсия
в отдельной молекуле
строго запрещена в
дипольном приближении, переходы
между ними в конденсированной среде и в растворах могут инициировать локальные
электромагнитные поля и спин-спиновое взаимодействие ядер, возникающие при
столкновении молекул воды друг с другом или в контакте с парамагнитными
центрами других молекул. В процессе столкновения двух молекул воды импульс
Т-фотона одной молекулы преобразуются в
момент импульса другой. Отметим, что в орто-изомере легче всего будут
возбуждаться вращательное состояние, отвечающее минимальному значению момента
инерции (J1), при этом вращению вокруг оси Х, в принципе, может
способствовать вращательный момент ядерных спинов протонов (Рис 7).
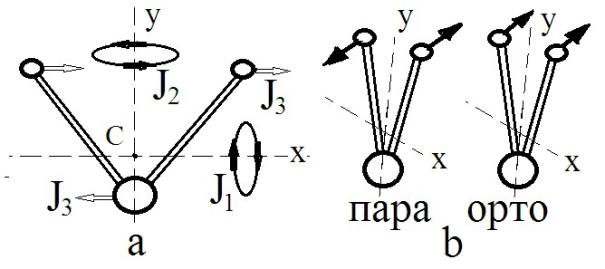
Рис 7. Схемы вращений
молекулы воды и ориентаций спинов протонов: а – вращения молекулы относительно
центра тяжести (С), отвечающие трем значениям моментов инерции (J1 = 1,02 10–47 кг м2; J2 = 1,92 10–47 кг
м2; J3 = 2,94 10–47 кг м2 [20]); b –
варианты ориентации ядерных спинов протонов в пара- и орто-изомере молекулы
воды.
В силу отсутствия вращения пара-изомер с большей
вероятностью будет образовывать водородные связи. Кинетика разрыва водородной
связи, очевидно, будет зависеть от энергии вращательного возбуждения молекулы,
которая в свою очередь пропорциональна ее моменту инерции. Если предположить,
что частоты вращения молекулы для всех трех J различаются не значительно, то время жизни среднестатистической водородной связи будет
иметь три значения, относящиеся между собой как экспоненты от J1 , J2, J3. Для значений J на Рис 7, эти отношения получаются 1 : 6,5 : 18.
Теоретические оценки времени жизни водородной связи дают величины 25, 125 и 500
фс [22]. Отношения между ними качественно согласуются с нашими оценками.
Селективное вращательное возбуждение молекул воды,
приводящее к разрыву водородных связей
и последующей ее ориентации в кулоновском поле канала аквапорина, лежит в
основе механизма работы водных каналов мембран (Рис 8). Можно предположить, что
в электромагнитном поле внутри белка поступательное движение молекулы воды сопровождается
ее вращением по принципу «пропеллера» (вращательная диффузия). Для выявления
селективности работы канала по отношению к орто/пара изомерам необходимо
определить его величину внутри клетки. Работа данного канала за счет
осмотических сил регулируется гормоном вазопрессином, который содержится в
крови и поддерживает тонус кровеносных сосудов.
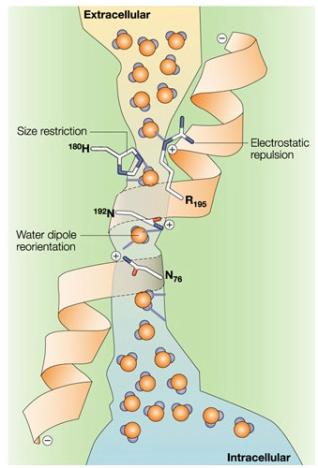
Рис 8. Схема работы водного канала мембраны,
образованного белком аквапорином. В центре канала внутримолекулярное кулоновское
поле меняет знак, что вынуждает диполи воды переворачиваться [27].
Эффекты квантовой реологии позволяют объяснить высокую
скорость прохождения эритроцитов по кровеносным капиллярам, размеры которых,
как правило, меньше самих эритроцитов. Установлено, что они при этом сильно
деформируются, сбрасывая из себя до половины находящейся в них воды по
аквапориновым каналам.
Считается [23], что деформируемость эритроцитов
является решающим фактором в уменьшении динамической вязкости крови. Прямыми
микроскопическими наблюдениями с кинорегистрацией было показано (Рис 9), что с
увеличением давления растет скорость кровотока по стеклянному капилляру и при
этом вблизи стенки наблюдается тем большая деформация эритроцитов, чем выше
скорость кровотока. При отсутствии движения эритроциты у стенки имеют круглую
форму, а при скорости 6 мм/с они принимают вытянутую форму в виде веретена.
Боковые поверхности эритроцита в капилляре располагаются касательно к слоям
разных скоростей, то есть вдоль оси сосуда. При этом достигается максимальная
эффективность диффузного обмена газами и метаболитами между кровью и межклеточной
жидкостью.
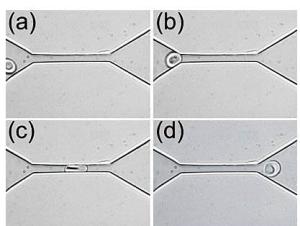
Рис 9. Механизм прохождения эритроцита диаметром ~7
мкм через капилляр диаметром ~4 мкм [23].
Эти данные дополняют исследования
зависимости вязкости крови от концентрации эритроцитов в сопоставлении их с зависимостью
отношения орто/пара изомеров воды от температуры (Рис 10).
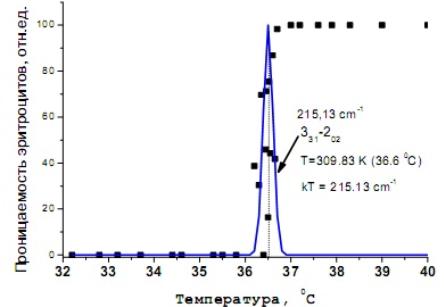
Рис 10. Зависимость
проницаемости (текучести) эритроцитов от температуры (квадратные точки) и
контур резонансной линии перехода пара-изомера воды с энергией 215 см–1
(Т = 309К) – синяя линия [21].
В работе [21] установили точное совпадение значения
тепловой энергии, при которой наблюдается
скачок «текучести» эритроцитов с
энергией кванта вращательного перехода в параизомере воды. Исходя из этого, предположили, что
вязкость крови при температуре 36,6оС резко падает из-за
лавинообразного перехода параизомеров воды, выходящей из эритроцита, в
ортоизомеры. Поскольку при данной температуре эффективно заселяются
вращательные состояния ортоизомера, то
возрастает вероятность разрыва водородных связей, что в итоге и приводит
к снижению динамической вязкости крови.
Таким образом, процесс снижения вязкости крови в
капиллярах многофакторный – в нем сочетаются:
-
безбарьерная зависимость вязкости воды от температуры в критических точках, к которым
относится и температура 36,6оС [10];
-
резкое усиление взаимодействий молекул воды в условиях капилляра с форменными
элементами, белками крови [21] и со стенками капилляра;
-
обогащение воды орто-изомерами и гомогенизация кластерно-комковой фазы [24].
В результате действия этих факторов возрастает
вращательно-поступательная подвижность молекул воды, что облегчает диффузию
газов и метаболитов через поры и стенки капилляров.
5. ХИРАЛЬНО-КОГЕРЕНТНЫЕ АНСАМБЛИ
Однородность сплошных сред и механизмы
пространственно-временной корреляции их элементов, превращает их в когерентные
ансамбли, играющие ключевую роль в
процессах адаптации и эволюции организма по стреле времени. Максимальную
чувствительность к внешним физическим факторам, включая ХФП, будут иметь
жидкостные системы (кровь, ликвор, синовия, глазная жидкость). Можно полагать
участие воды в механизмах чувствительности к ХФП и других сплошных тканей. К
примеру, в состав подкожной клетчатки содержит 70% в той или иной степени
связанной воды, паренхима органов до 90% и костная ткань до 10%. Конечно, у
этих тканей будут свои энергетические диапазоны чувствительности к ХФП. Общие
физико-химические принципы действия и типы когерентных биосистем организма даны
в [5, 10, 16, 19]. Их можно уточнить, используя данные по хиральной гистологии
и квантовой реологии тканей.
В биосистемах источником Т-фотонов и ЭМ-ЭФ являются
ускоренные движения электрона и других заряженных частиц. Соответственно,
действию квантов ЭМ-энергии подвержен только химический уровень структурной
организации ансамблей. В рамках ахиральной ЭМ-энергетики осуществляются
трофическая и репродуктивная функции любой живой твари, включая человека.
Хиральность ЭМ-энергетики, связанная с
ФАМ, у неразумных тварей может возникнуть лишь в патологической форме
как следствие извращения метаболизма под действием аномального внешнего фактора
[5].
Разумность же человека, напротив, является
закономерным следствием развития чувствительности его метаболизма к действию универсального
ХФП в процессе эволюции. На ранних этапах эволюции под непрерывным действием
солнечных нейтринных Х-ЭФ на генеалогическом древе жизни образовалась хиральная
ветвь, плодом которой в итоге явился homo sapiens. Его способность мыслить,
то есть созидать новый смысл, находится в прямой зависимости от степени
хиральности его мозга (ФАМ) и эффективности подпитки организма хиральной
энергией Х-ЭФ за счет действия хирально-когерентных ансамблей. Интенсивность их
работы определяются плотностью потока Х-ЭФ в приземном эфире [27] и уровнем
умственно-духовного развития человека [25]. Оба эти фактора зависят от геокосмических
условий [26].
Нейтрино (антинейтрино) является продуктом
неравновесной динамики нуклонных или субъядерных элементов и образуется в
космосе, а также в результате реакций бета-распада радиоактивных ядер или
свободного нейтрона [25]. Стационарный поток солнечного нейтрино модулируется
воздействием на энергетику Солнца магнитного поля планет и Юпитера главным образом
[9]. Солнечное нейтрино нестабильно и
рассеивается в межпланетном магнитном поле на N квантов Х-ЭФ изоморфных нейтрино, ими и насыщается
метрика приземного эфира [25, 27]. В здоровом организме стационарных экзогенных
источников Х-ЭФ быть не может, главным образом из-за деструктивного действия на
ткани сопутствующих нейтрино продуктов бета-распада.
Учитывая ядерный генезис нейтрино и Х-ЭФ, предположили
[5], что их поглощение-конденсация в биосистеме возможны в коррелированных
ансамблях спинов ядер или их магнитных моментов. Ориентация отдельных ядерных
спинов в ансамблях будет зависеть от сверхтонких спин-спиновых и
спин-орбитальных взаимодействий ядер и электронов, а степень спиновой
когерентности всего ансамбля будет модулироваться магнитными полями как
внутренними, так и внешними. Ориентация орбитального момента электрона определяется
геометрией молекулярных орбиталей и ориентацией молекулы, что может обусловить
зависимость чувствительности ансамбля ядерных спинов к ХФП от степени упорядоченности
атомов и молекул. Вклад во вращательную энергию молекулы воды ядерного спина
составит 0,1 – 1 Дж/моль [12], а энергия его нутации и прецессии, очевидно,
будет меньше еще на один-два порядка.
Можно предположить, что через влияние на параметры этих движений спина
протона кванты Х-ЭФ будут способствовать самоорганизации когерентных ансамблей.
Учитывая присутствие воды во всех тканях организма,
взаимодействия Х-ЭФ с протонами можно взять за основу механизма
чувствительности когерентных ансамблей к ХФП. Дополнительным ориентирующим
фактором можно считать кулоновские поля положительно или отрицательно
заряженных регулярных центров на белковых молекулах, полисахаридах или на поверхностях
клеток и капилляров, а также в структурах соединительной и костной ткани [26].
В упорядоченных таким образом гидратных оболочках или цепочках по схеме (1)
будет идти конденсация квантов Х-ЭФ с передачей их на хиральные центры
гидратации, что равносильно их активации. Отметим, что процесс конденсации
нейтринной энергии в жидкостных системах организма будет идти главным образом в
состоянии сна, когда снижается температура организма на градус [16, 25].
Возможные механизмы включения квантов ЭМ-ЭФ и Х-ЭФ в
метаболизм мозга по гуморальным каналам (кровь, глазная жидкость), обсужден в
[16, 27]. К ним можно добавить существенную роль аквапориновых каналов (Рис 8)
в регуляции водного баланса мозга, активности нейроглий и внеклеточного К+
клиренса [28]. Данные каналы, по-видимому, лимитируют кинетику образования
ликвора и метаболизм третьего желудочка. Учитывая вращательный характер
диффузии воды через аквапориновые каналы, они могут играть роль хиральных
фильтров, контролирующих асимметрию биоэнергетики ликвора и процесса обмотки
аксонов миелиновой оболочкой.
Вегетативная нервная система (ВНС) вносит свой вклад в
процесс акцептирования и передачи в мозг хиральных квантов ЭМ-Ф и Х-ЭФ [19].
Они могут конденсироваться в процессе ночного сна, например, в подкожной
клетчатке на нервных окончаниях и в капиллярных анастомозах. После пробуждения в процессе физических
упражнений данные кванты, очевидно, асимметрично активируют ВНС, что в
совокупности с асимметрией речевой функции составляет основу ресурса ФАМ, который
проявляется при беге на месте [16, 19]. Конденсат правых Х-ЭФ может
накапливаться в нервных окончаниях подкожной клетчатки, соединительных тканей и
мышц во сне или в состоянии длительной неподвижности тела, вследствие чего
возникает рефлекторное желание потянуться, при этом деформация тканей порождает
эфферентные импульсы, которые задействуют в мозгу центры удовольствия.
Хиральные метаболиты, очевидно, не только участвуют в
формировании миелиновых оболочек аксонов с определенным знаком спирали, но и в
активации психических и когнитивных функций мозга. Например, при некоторых
психосоматических заболеваниях (шизофрения, болезнь Альцгеймера и Паркинсона),
наблюдаются значительные изменения уровня некоторых D-аминокислот в плазме
крови, сером и белом веществе головного мозга, спинномозговой жидкости [29]. С
этими данными можно сравнить эффект действия хиральных лекарств [30].
Обнаружено [19], что знак ресурса ФАМ может инвертировать, если в течение
предшествующего дня и на ночь принимается лекарство, имеющее, например,
выраженное действие на дыхательную функцию. Кроме того, ресурс ФАМ у праворуких
мужчин и женщин имеет противоположные знаки.
ЗКЛЮЧЕНИЕ
Настоящий обзор показал, что в физиологии человека на
уровне функций и морфологии существенную роль играют оптически активные
метаболиты и хиральные среды, обладающие кооперативными свойствами и
чувствительностью к внешним физическим факторам. Установлено, что для
объяснения механизмов влияния экзо- и эндогенных хиральных факторов на метаболизм
и психофизиологию человека необходимо привлекать квантово-механические и
квантово-химические свойства отдельных молекул и когерентных ансамблей.
Предложенный в работе математический язык квантового морфогенеза, в принципе,
показал свою работоспособность. Можно надеяться, что данный язык и высказанные
гипотезы о механизме влияния хирального фактора на морфогенез и духовную
эволюцию человека, получат в дальнейшем экспериментальное подтверждение и
послужат во благо развития науки о человеке.
ЛИТЕРАТУРА
1.Холманский А.С. Энергоформа //http://www.sciteclibrary.ru/rus/catalog/pages/7441.html; Фрактально-резонансный принцип действия // www.chaos-trading-group.narod.ru/chaos.htm
2. Берг Л.С. Номогенез, или эволюция на основе закономерностей, Гл. 5 в кн. Теория Эволюции, 1922 // http://www.evolbiol.ru/berg.htm
3. Казютинский В.В., Балашов Ю.В. Антропный принцип // Природа, 1, 1989, http://vivovoco.rsl.ru/VV/JOURNAL/NATURE/OLD/ANTROP.HTM
4. Холманский А.С. Электромагнитная природа релятивистских эффектов // http://sgma.alpha-design.ru/MMORPH/N-19-html/kholmanskiy-2/khomanskiy-2l.htm; Апофеоз ахиральности // http://www.sciteclibrary.ru/rus/catalog/pages/9123.html; Хиральная пустота // http://www.sciteclibrary.ru/rus/catalog/pages/9091.html/t_blank
5. Холманский А.С. Адаптация растений к аномальным физическим факторам. http://www.smolensk.ru/user/sgma/MMORPH/N-23-html/holmanskiy/holmanskiy.htm; Зависимость ресурса функциональной асимметрии мозга от внешних условий // http://www.j-asymmetry.com/Kholmansky_1_09.htm
6. Холманский А.С. Модель одухотворенного мироздания // www.sciteclibrary.ru/rus/catalog/pages/8084.html; Духовно-физический изоморфизм // www.sciteclibrary.ru/rus/catalog/pages/9069.html;
7. Верещагин И.А. Корона Гамова венчает физику ХХ века // Успехи современного естествознания, -2006. -№ 8. –С. 29;
http://www.rae.ru/use/?section=content&op=show_article&article_id=4135
8. Клапдор-Клайнгротхаус Г. В., Штаудт А. Неускорительная физика элементарных частиц. М. 1997. 527 с.
9. Холманский А.С.// Начала православной науки
http://library.by/portalus/modules/psychology/readme.php?subaction=showfull&id=1132581314&archive=01&start_from=&ucat=1&; Теофизика солнца //
http://www.quantmagic.narod.ru/volumes/VOL422007/p2209.html
10. Холманский А.С. Особенности термодинамических свойств воды и биоэнергетика // Доклады РАСХН. №2. 2006. С. 63; http://www.sciteclibrary.ru/rus/catalog/pages/7897.html; Зависимость от температуры оптической активности физиологических растворов сахаров // Математическая морфология. 2006.
http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky/holmansky.htm
11. Бункин А.Ф. Нурматов А.А., Першин С.М. Когерентная четырехфотонная спектроскопия низкочастотных либраций молекул в жидкости // УФН, -2006. –Т.176. -№8. –С. 883-889
12. Кизель В.А. Практическая молекулярная спектроскопия. М.: -1998. – 254 с.
13. Белоусов Л.В., Войеков В.Л., Попп Ф.А., Митогенетические лучи Гурвича: драматическая история и новые перспективы // Природа, - 1997. -№ 3. С. 64—80.
14. Дайсон Ф., Монтропл Э., Кац М., Фишер М., Устойчивость и фазовые переходы, М. «Мир», -1973; . Сент-Дьердьи А. Биоэнергетика, -М.: -1960. -156 с.
15. АТФ-синтаза // http://ru.wikipedia.org/wiki/АТФ-синтаза
16. Холманский А.С. Ресурс функциональной асимметрии мозга // Математическая морфология. 2006; http://www.smolensk.ru/user/sgma/MMORPH/N-12-html/holmansky-4/holmansky-4.htm Моделирование физики мозга // Там же. 2006, - Т.5, - В.4. http://sgma.alpha-design.ru/MMORPH/N-12-html/holmansky-3/holmansky-3.htm; Сознание и физическая реальность. -2008. -№12. –С.23-38
17. Feldman Y, et al, Human skin as arrays of
helical antennas in the millimeter and submillimeter waverange //
Phys.Rev.Lett. -2008. –V.100. –P. 128102
18. Takagi
S., Tagawa M. Predominance of right-handed spirals in human eccrine sweat ducts // Japan.J.Physiol.
-1955. –V.5(2). –P. 122-130
19. Холманский А.С. Дихотомия правого и левого в живых системах // Асимметрия. -2008. –Т. 2. -№ 3. –С. 60-67. www.j-asymmetry.com/Asymmetry_03_2008.pdf ; Холманский А.С., Минахин А.А. Морфологические и физические факторы асимметрии вегетативной нервной системы человека // Всероссийская конференция «Современные направления в исследовании функциональной межполушарной асимметрии и пластичности мозга». -М. -2010. -С.270-274.
20. Еремина В.В. Сравнительный анализ моментов инерции свободной и закрепленной молекулы воды // Кибернетическая физика. 2009. №1 (19); http://www.amursu.ru/attachments/ics/N19_07.pdf
21. Першин С.М. Орто/пара конверсия Н2О в воде и скачок «текучести» эритроцитов через микрокапилляр при температуре 36.6±0.3оС // Сборник избранных трудов V Международного конгресса «Слабые и сверхслабые поля и излучения в биологии и медицине». Санкт Петербург. -2009. –С.89-99
22. Волошин В.П., Наберухин Ю.И. Распределение времени жизни водородных связей в компьютерных моделях воды //Ж. структурной химии. -2009. –Т. 50, -N. 1 С. 84 – 95
23. Чернух А.М., Александров П.Н., Алексеев О.В.
Микроциркуляция. М.: -1975.
456 с.
24. Huang
C., Wikfeldt K.T., Tokushima T., et al.
The inhomogeneous structure of water at ambient conditions. Proceedings of the
National Academy of Sciences; http://www.pnas.org/content/early/2009/08/13/0904743106
25. Холманский А.С. Галактический фактор духовной эволюции // http://www.j-asymmetry.com/Kholmansky_1_09_2.htm; Ядерно-химический катализ // http://www.sciteclibrary.ru/rus/catalog/pages/6303.html; Теофизика нейтрино // Квантовая магия. -2007. –Т.4. –В.2. http://quantmagic.narod.ru/volumes/VOL422007/p2148.html
26. Холманский А.С., Минахин А.А., Дегтярев В.П. Модели и аналогии в физиологии зубов // Математическая морфология. - Т. 9. -В. 3. - 2010.
http://www.smolensk.ru/user/sgma/MMORPH/N-27-html/kholmanskiy/kholmanskiy.htm
27. Холманский А.С. Мифология и физиология алкоголя // Сознание и физическая реальность. -2009. -№7. http://www.smolensk.ru/user/sgma/MMORPH/N-21-html/holmansky-1/holmansky-1.htm; Метрика приземного эфира // http://www.sciteclibrary.ru/rus/catalog/pages/7876.html
28. Amiry-Moghaddam M.,
Ottersen Ole P. The molecular basis of
water transport in the brain // Nature Reviews Neuroscience. -2003. -V.4. -№ 12.
P. 991-1001 http://mglinets.narod.ru/slova2/brnWater.htm
29. Червяков А.В. Нарушение молекулярной асимметрии аминокислот (D\L-энантиомеры) при нормальном старении и нейродегенеративных заболеваниях // Асимметрия. -2010. -Т. 2; http://www.j-asymmetry.com/Chervyakov_2_2010.htm
30. Твердислов В.А., Яковенко Л.В., Жаворонков А.А. Хиральность как проблема биохимической физики // Рос. Хим. Журн., -2007, -Т. LI. -№ 1, С. 13-22.
CHIRALITY AND QUANTUM
EFFECTS AS FACTORS MORPHOGENESIS
Kholmanskiy A.S.
The
review is devoted a role substantiation chiral dichotomi in world evolution on
an arrow of time from simple to the difficult. Are presented axiomatics and
rules of application of the universal mathematical language allowing uniformly
to describe law of change of discrete forms of a matter and quantum conditions
of biosystems at all levels of their organization from subelementary to socially-spiritual.
The analysis chiral the anatomic and functional elements of human physiology
forming quantum coherent ensembles, sensitive to external physical factors
chirality, as electromagnetic, and neutrinos the nature is carried out. The
role of quantum properties of water in bio-energetic and processes of
self-organizing of coherent ensembles is analyzed. Being guided by logic
antrope a principle, an evolution vector have identified with a gradient
chirality and have connected with it degree chiral cleanliness of a brain with
which asymmetry of a brain and efficiency of heuristic thinking have correlated
functional.
Key
words: biosystem, a morphogenesis, energoform,
quantum, asymmetry, nuclear spin, water, a brain.
Московский государственный университет инженерной экологии (МГУИЭ)
Поступила в
редакцию 17.12.2010.