
УДК 612.833.81
К 100-летию со дня
рождения академика П. К. Анохина
© 1997 г.
В. А. Правдивцев, С. Б. Козлов
27 января 1998 году исполнилось 100 лет со дня рождения выдающегося нейрофизиолога ХХ столетия академика АН СССР, академика АМН СССР Петра Кузьмича Анохина. Имя Петра Кузьмича Анохина, основоположника теоретической физиологии, физиологической кибернетики, автора глобальной концепции функциональной системы как единицы целостной интеграции, стоит в одном ряду с именами Р. Декарта, И. М. Сеченова, И. П. Павлова.
Неоднократно научная деятельность П. К. Анохина пересекалась со Смоленском. Так, еще в 30 годы в аспирантуру к П. К. Анохину поступает ассистент кафедры физиологии медицинского факультета Смоленского университета Иосиф Иванович Лаптев. В лаборатории П. К. Анохина он успешно впервые в мире изучал ЭЭГ-компоненты условного рефлекса. В пятидесятые годы в Смоленской областной типографии бы отпечатана одна из наиболее известных монографий П. К. Анохина - “ Внутренне торможение как проблема физиологии”. 11 лет (1958-1969 гг.) кафедру нормальной физиологии Смоленского медицинского института возглавлял ученик П. К. Анохина профессор Яков Андреевич Милягин. Профессор Я. А. Милягин заложил основу творческих научных контактов между кафедрой нормальной физиологии Смоленского медицинского института и кафедрой нормальной физиологии 1 Московского медицинского института, которой заведовал академик П. К. Анохин.
Юбилейные мероприятия, посвященные
100-летию со дня рождения академика П. К. Анохина,
прошли в Москве 27-28 января 1998 г.
ОСНОВНЫЕ ДАТЫ ЖИЗНИ И ДЕЯТЕЛЬНОСТИ АКАДЕМИКА
П. К. АНОХИНА
1898 г. 14 (27) января Родился в г. Царицыне.
1913 г. Окончил Царицынское городское училище.
1915-1918 гг. Студент Новочеркасского землемерно-агрономического училища.
1918-1919 гг. Доброволец Доно-Ставропольского партизанского отряда на северном участке Царицынского фронта, старший топограф штаба 10-ой Красной Армии, инспектор штаба обороны города по возведению укреплений.
1920-1921 гг. Член Исполкома горсовета г. Новочеркасска, комиссар по печати, заведующий Центропечатью Донского округа, ответственный редактор газеты "Красный Дон" (Новочеркаск).
1921-1926 гг. Студент Ленинградского государственного института медицинских знаний (ГИМЗ). Под руководством В. М. Бехтерева на 1-ом курсе проводит первую научную работу "Влияние мажорных и минорных колебаний звуков на возбуждение и торможение в коре головного мозга".
1922-1930 гг. В Военно-медицинской академии, а затем в Институте физиологии АН СССР проводит исследования под руководством И. П. Павлова по изучению процессов, протекающих в корковом центре условного раздражителя во время применения безусловного сигнала, а также работы по изучению природы внешнего и внутреннего торможения.
1926 г. Окончил Государственный институт медицинских знаний и получил специальность врача.
1926-1930 гг. Старший ассистент, доцент кафедры физиологии Ленинградского Зоотехнического института. Продолжал работать в Павловской лаборатории. На кафедре под руководством учеников Н. Е. Введенского - профессоров Ф. Е. Тура и Ю. М. Уфлянда выполнил исследования по изучению особенностей кровоснабжения головного мозга, роли кровяного давления для активного сокращения капилляров, влияния ацетилхолина на сосудистую и секреторную функции слюнной железы.
1930 г. По рекомендации И. П. Павлова избран профессором кафедры физиологии медицинского факультета Нижегородского университета (впоследствии Горьковского медицинского института).
1930-1935 гг. Заведующий кафедрой физиологии Горьковского медицинского института и кафедрой физиологии биологического факультета Нижегородского университета. Предложил принципиально новые методы изучения условных рефлексов: секреторно-двигательный метод, позволявший изучать не только классический секреторный компонент, но и двигательную реакцию животного в динамике условного рефлекса, а также оригинальный метод с внезапной подменой безусловного подкрепления. Этот метод позволил П. К. Анохину прийти к заключению о формировании в центральной нервной системе специального аппарата, в котором заложены параметры будущего подкрепления (своего рода модель будущего результата "заготовленное возбуждение"). Позже этот аппарат получил название "акцептор результата действия". В это же время вводится понятие "санционирующая афферентация", позже "обратная афферентация". Приступил к исследованиям по изучению центрально-периферических отношений в нервной деятельности с помощью метода гетерогенных анастомозов нервов. В предисловии к коллективной монографии "Проблемы центра и периферии в физиологии нервной деятельности" (1935) дает первое определение "функциональной системы". Создает и руководит Горьковским отделением Всесоюзного общества физиологов. биохимиков и фармакологов.
1933-1935 гг. Директор Отделения эволюционной физиологии высшей нервной деятельности Горьковского филиала Всесоюзного института экспериментальной медицины (ВИЭМ), где развернул работы по онтогенезу высшей нервной деятельности.
1934 г. Кафедра физиологии Горьковского медицинского института, руководимая П. К. Анохиным, занимает первое место на Всесоюзном конкурсе ВУЗов.
1935 г. П. К. Анохину присуждена ученая степень доктора медицинских наук. Принял участие в работе ХУ Международного физиологического конгресса в Москве, где выступил с докладом "Единство центра и периферии".
1935-1944 гг. Заведующий отделом нейрофизиологии ВИЭМ. В отделе продолжил углубленные исследования проблемы центра и периферии в нервной деятельности не только в общебиологическом, но и в эмбриологическом аспекте.
1936-1942 гг. Профессор кафедры физиологии и патологии высшей нервной деятельности Центрального института усовершенствования врачей-(ЦИУ) Москва.
1938-1945 гг. Заведующий психоневрологическим сектором (с 1942 г. физиологическая лаборатория) Центрального нейрохирургического института Академии медицинских наук СССР (Москва) - исследовал физиологические свойства нервного рубца и влияния на него физиотерапевтических факторов. Значительное место в клинико-физиологических исследованиях П. К. Анохина занимают работы по изучению травм периферической нервной системы. Создал новый метод замещения дефектов нервного ствола с помощью формализированного нерва.
1941-1942 гг. Эвакуирован в Томск, где руководит нейрохирургическим отделением травм периферической нервной системы эвакогоспиталя. Внимание П. К. Анохина и его сотрудников в этот период сосредоточено на изучении военных травм периферической нервной системы, механизмов регенеративных процессов, обеспечивающих восстановление анатомической целостности и физиологических функций травмированных нервных стволов, изучает механизм возникновения каузалгических болей. Предложил метод их предупреждения, сформулировал "физиологическую теорию нервного рубца". Активно внедряет в повседневную хирургическую практику метод трансплантации формалинизированного нерва. Разработал и внедрил в медицинскую практику новые электрофизиологические методы диагностики военных травм нервных стволов : метод характеристики "объема движений" и метод градуированной частоты раздражения. Все эти работы нашли отражение в монографии "Пластика нервов при военной травме периферической нервной системы" (1944).
1942-1945 гг. Профессор кафедры физиологии биологического факультета Московского университета им. Ломоносова.
1943 г. За самоотверженную работу в эвакогоспиталях по лечению бойцов и командиров Красной Армии, раненых в боях с немецкими захватчиками, успешное проведение противоэпидемических мероприятий, хорошую организацию медицинского обслуживания населения и подготовку медицинских кадров награжден орденом "Знак почета".
1944-1950 гг. Заведующий Отделом физиологии нервной системы Института физиологии АМН СССР, с 1946 г. -заместитель директора Института по научной работе, с 1949 г. - директор Института.
1945 г. Избран действительным членом Академии медицинских наук СССР.
1945-1946 гг. Начальник секретариата Президиума АМН СССР. Опубликовал монографию "от Декарта до Павлова".
1946 г. Председатель научно-плановой комиссии АМН СССР 1946-1950 г. г. Член Президиума АМН СССР.
1950 г. На научной сессии, посвященной проблемам физиологического учения академика Павлова, развиваемое П.К. Анохиным направление тендециозно объявлено ошибочным. Отмечалось, что П. К. Анохин "не двигал вперед павловское учение и на протяжении многих лет занимался ревизией идейных основ материалистического учения Павлова о высшей нервной деятельности, принижая его значение". П. К. Анохин был освобожден практически от всех постов и лишен возможности проводить научные исследования в Москве.
1950-1952 гг. Заведующий кафедрой физиологии Рязанского медицинского института.
1951-1958 гг. Заведующий лабораторией физиологии и патологии Института хирургии им. А. В. Вишневского АМН СССР. В лаборатории продолжил исследования по высшей нервной деятельности, патогенезу гипертонической болезни, исследовал вопросы гипотермии, роль ретикулярной формации в механизме наркоза, болевых ощущений и т.д. Работы по изучению закономерностей компенсации нарушенных функций обобщил в брошюре "Общие принципы компенсации нарушенных функций и их физиологическое обоснование" (1955).
1953-1955 гг. Профессор, заведующий кафедрой физиологии и патологии высшей нервной деятельности ЦИУ врачей.
1955-1974 гг. Заведующий кафедрой нормальной физиологии 1-го Московского ордена Ленина и ордена Трудового Красного Знамени медицинского института им. И. М. Сеченова. Руководитель Сеченовского института физиологии. В этот период разрабатывает и окончательно формулирует основные положения теории функциональных систем, включает в операциональную архитектонику функциональной системы ряд основных узловых механизмов, развертывающихся в определенной динамической последовательности: афферентный синтез, стадию принятия решения, акцептор результата действия, программу действия, результат деятельности, обратную афферентацию от результата действия. На основе теории функциональных систем разработал теорию системогенеза. Предложил новую трактовку роли ретикулярной формации в механизмах сна и бодрствования, механизмах боли и избирательного действия наркотических средств, нейрохимическую трактовку различных компонентов вызванного потенциала, экспериментально обосновал конвергентную теорию замыкания условного рефлекса, предложил гипотезу интегративной деятельности нейрона и др.
1956-1974 гг. Член редколлегии, а с 1965 г. - член редакционного совета "Физиологического журнала им. И. М. Сеченова".
1959-1974 гг. Заведующий лабораторией общей физиологии центральной нервной системы Института нормальной и патологической физиологии АМН СССР.
1960-1974 гг. Редактор раздела "Физиология" 2-го издания Большой медицинской энциклопедии (БМЭ), член редакционной коллегии З издания БМЭ.
1961 г. Награжден орденом Трудового Красного Знамени за "большие заслуги в подготовке специалистов и развитии науки".
1962-1974 гг. Член редколлегии "Журнала высшей нервной деятельности им. И. П. Павлова".
1965 г. Награжден Почетной грамотой Советского комитета защиты мира за активную деятельность по укреплению мира между народами.
1966 г. Избран действительным членом (академиком) Академии наук СССР. Награжден орденом Трудового Красного Знамени "за большие заслуги в области охраны здоровья советского народа и в развитии медицинской науки и медицинской промышленности".
1966-1974 гг. Член бюро Отделения физиологии АН СССР.
1968 г. Награжден Золотой медалью им. И. П. Павлова за серию работ по физиологии центральной нервной системы, разработку нового направления современной нейрофизиологии - системного подхода в изучении функциональной организации мозга. На Международном конгрессе физиологов в США выступил с докладом "Конвергенция возбуждений на нейроне как основа интегративной деятельности мозга".
1968-1971 гг. Заместитель председателя Объединенного научного совета "Физиология человека и животных" АН СССР.
1969-1974 гг. Член редколлегии журнала "Нейрофизиология".
1970-1974 гг. Председатель Московского физиологического общества. Председатель Комиссии по документальному наследию академика И. П. Павлова АН СССР.
1971-1974 гг. Главный редактор журнала "Успехи физиологических наук",
1972 г. Присуждена Ленинская премия за монографию "Биология и нейрофизиология условного рефлекса", опубликованную в 1968 г. На Международном конгрессе психологов в Японии выступил с докладом "Память как компонент системной деятельности мозга".
1974 г. Публикация работы "Системный анализ интегративной деятельности нейрона", где были сформулированы основные идеи о внутриней-рональной переработке информации.
5 марта Скончался в Москве, похоронен на
Новодевичьем кладбище.
ПОЧЕТНЫЕ ЗВАНИЯ П. К. АНОХИНА
Действительный член Венгерской академии наук. Почетный доктор наук Будапештского государственного университета им. Л.Этвеша.
Почетный доктор Лейпцигского университета. Почетный член Чехословацкого медицинского общества им. Я. Е. Пуркинье.
Член Международной организации по
исследованию мозга. Член Национального
географического общества США.
Член-корреспондент Общества биологической
психиатрии США. Член Павловского научного
общества США.
ПАМЯТИ ВЫДАЮЩЕГОСЯ УЧЕНОГО
В 1974 г. в честь П. К. Анохина учреждена
премия им. академика П. К. Анохина АМН СССР за
лучшую научную работу по нормальной физиологии.
В Московской медицинской академии им. И. М.
Сеченова учреждена именная стипендия Ученого
Совета имени П. К. Анохина для студентов за
выдающиеся успехи в учебе и исследовательской
деятельности. С 1976 г. проводятся научные
"Анохинские чтения", целью которых является
ознакомление общественности о ходе развития
научного наследия ученого. Имя академика П. К.
Анохина присвоено Научно-исследовательскому
институту нормальной физиологии РАМН, а также
одной из улиц Москвы. В 1992 г. в
Научно-исследовательском центре "Медицинский
музей РАМН" создан мемориальный кабинет П. К.
Анохина. В Москве, Нижнем Новгороде и Рязани в
память о П. К. Анохине установлены мемориальные
доски.
Изучение феномена условного рефлекса развивается по нескольким направлениям. Первое - связано с анализом нейрофизиологической природы временной связи. Вместе с тем, не менее важным является второе направление, в рамках которого исследуется принципиальная архитектура, общая организация условнорефлекторного поведенческого акта.
Как трактовалась общая организация условнорефлекторного акта на начальном этапе его изучения? Условнорефлекторный акт рассматривался с позиций классической рефлекторной теории. Достаточно обратить внимание на название, которое было предложено для обозначения нового приобретенного в ходе обучения поведенческого акта - условный рефлекс, под которым понимали ответную рефлекторную реакцию, возникающую при определенных условиях. Принципиальная схема условнорефлекторного поведенческого акта представлялась в соответствии с рис. 1.
На схеме видно, что условный сигнал, взаимодействуя с некоторыми фоновыми раздражителями, запускает через ЦНС реакцию, проявляющуюся в активности периферических эффекторов 1, 2 и т. д. С точки зрения современных представлений такого рода подход к пониманию архитектуры условного рефлекса является упрощенным, скрывающим истинную сущность поведенческого акта как процесса физиологической саморегуляции.
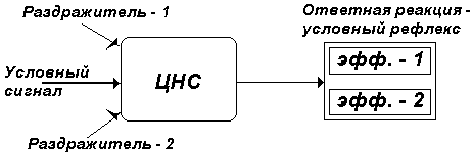
Рис. 1. Схема общей организации
условнорефлекторного поведенческого акта с
позиций рефлекторной теории. Описание в тексте.
Анализ механизмов саморегуляции
жизненно важных констант организма (кровяное
давление, напряжение углекислого газа и
кислорода в артериальной крови, температура
внутренней среды, осмотическое давление плазмы
крови, стабилизация центра тяжести в площади
опоры и т. д.) показывает, что аппаратом
саморегуляции выступает функциональная
система (П. К. Анохин), принципиальная
структура которой является универсальной (рис. 2).
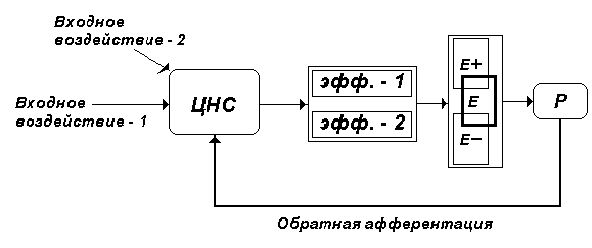
Рис. 2. Принципиальная схема
функциональной системы как аппарата
саморегуляции (по П. К. Анохину). Е - конечный
полезный результат функциональной системы. Е-, Е+
- отклонения конечного полезного результата под
влиянием различных факторов от оптимального
значения (Е). Р - рецептор функциональной системы,
точно приспособленный к свойствам полезного
результата. ЦНС - центральная нервная система,
осуществляющая синтез всех входных воздействий.
ЭФФ.-1, ЭФФ.-2 - эффекторы в составе функциональной
системы, активность которых определяет
конкретный полезный результат.
Центральным пунктом функциональной системы выступает конечный полезный результат, определяющий динамическую активность всех ее узлов вследствие непрерывной передачи информации по каналам обратной связи (обратной афферентации) в ЦНС.
Рассмотрим архитектуру условнорефлекторного пищедобывательного поведенческого акта с позиций теории функциональной системы П. К. Анохина.
Результаты многочисленных опытов, проведенных в лаборатории П.К. Анохина, позволяют заключить, что иницативная роль в формировании поведенческого акта принадлежит исходным потребностям организма. Применительно к условнорефлекторному пищедобывательному поведенческому акту речь идет о потребностях в питательных веществах, расходуемых в процессе жизнедеятельности, что приводит к уменьшению их концентрации в циркулирующей крови и, как следствие, к активации специализированного центра голода, расположенного в гипоталамической области (рис. 3).
По множественным путям возбуждение от центра голода распространяется на различные отделы головного мозга, включая кору больших полушарий, при этом формируется субъективно неприятное чувство голода.
Эмоционально окрашенное состояние, возникающее как результат объективной потребности организма в питательных веществах, избирательно объединяющее нервные элементы различных уровней мозга, получило название пищевой мотивации, или мотивации голода (К. В. Судаков).
Экспериментальные данные показывают,
что именно мотивационные возбуждения в
каждом случае формируют основу той или иной
специфической формы целенаправленного
поведения, ориентированного на достижение в
среде обитания необходимого полезного
результата, устраняющего причины возникновения
первичного мотивационного возбуждения (рис. 4).
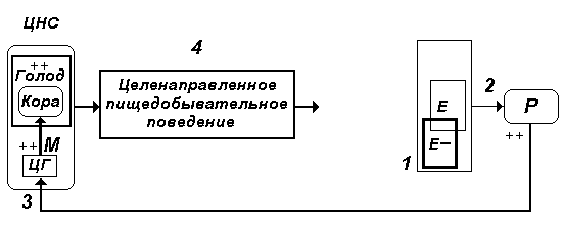
Рис. 3. Механизм инициирования
целенаправленного пищедобывательного поведения
на основе мотивационных возбуждений, отражающих
потребность организма в питательных веществах. 1.
Е- отклонение концентрации питательных веществ в
крови в ходе метаболизма от оптимального
значения Е. 2. Р- рецепторные приборы (центральные
и периферические), фиксирующие отклонение
параметра внутренней среды организма Е от
оптимального значения. При этом возникают
определенной интенсивности (++) импульсации,
активирующие гипоталамический центр голода - ЦГ
(3). На основе мотивационных возбуждений М (++)
происходит активация коры больших полушарий,
формируется чувство голода (++), инициирующее
целенаправленное пищедобывательное поведение -
4.
Как доказать, что в основе пищевых условных рефлексов лежит мотивация голода? Доказательство весьма простое и убедительное. Оказывается, если до опыта подопытное животное плотно накормить, условный пищевой рефлекс у такого животного не вырабатывается. Если накормить животное с выработанным условным пищевым рефлексом, в опыте условный рефлекс перестает проявляться. Таким образом, пищевой условнорефлекторный поведенческий акт следует рассматривать как пищедобывательный поведенческий акт, формирующийся на основе пищевой мотивации, или мотивации голода.
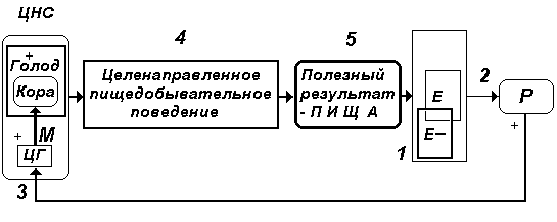
Рис. 4. Завершение процесса
пищедобывательного поведения достижением
конечного полезного результата (5) - нахождением в
среде обитания пищи и ее потреблением. В этом
случае повышается концентрация питательных
веществ в крови (1), уменьшается степень
возбуждения рецепторных приборов (2), уменьшается
активация центра голода (3), снижается
выраженность чувства голода. Цикл повторяется до
стабилизации параметра Е на оптимальном уровне.
Представленная на рис. 4 схема подчеркивает саморегуляторный характер пищедобывательного поведенческого акта, завершение которого связано с нахождением и потреблением в достаточном количестве пищи, обеспечивающей восстановление на оптимальном уровне концентрации питательных веществ в крови. Это приводит к изменению характера импульсаций рецепторных приборов функциональной системы, абсолютному угнетению центра голода, возбуждению гипоталамического центра насыщения, смене эмоционального состояния организма и прекращению пищедобывательной активности животного.
Рассмотрим более подробно содержание афферентных процессов, протекающих на уровне ЦНС и предопределяющих центральную организацию пищедобывательного поведенческого акта.
Мотивационные возбуждение, как отмечалось выше, создают базу для развертывания поведенческого акта. Вместе с тем, следует отметить определенную роль иных возбуждений, поступающих в ЦНС и активно влияющих на процесс центральной организации поведенческого акта.
Прежде всего, следует подчеркнуть необходимость учета обстановочных афферентаций (П. К. Анохин), под которыми понимают совокупность всех внешних факторов и идущих от них воздействий. Функциональное значение комплекса обстановочных возбуждений состоит в том, что они, наряду с мотивационными возбуждениями, дополнительно активируя аппараты памяти, оживляют центральные поведенческие программы, адекватные той обстановке, в которой реально пребывает организм, решая поведенческую задачу.
Приведем простые наблюдения, показывающие, как обстановочная афферентация влияет на организацию поведенческого акта. Например, голодная собака вблизи источника пищевого подкрепления обычно реализует поведенческую пищедобывательную программу. Однако, даже очень голодная собака, брошенная в воду, т. е. помещенная в необычную обстановку, в первую очередь запускает поведенческие программы, обеспечивающие процедуру спасения и выхода из воды, а не поиска пищи. П.К. Анохин, характеризуя значимость обстановочной афферентации для формирования адекватного типа поведения, часто приводил пример из клинической практики. Стандартный вопрос врача больному: “На что жалуетесь?” Безотносительно к обстановке ответ на поставленный вопрос может вызвать множество вариаций. На что человек жалуется в принципе? Однако, в обстановке поликлиники на уровне структур головного мозга субъекта “заранее создается такая интеграция нервных процессов, для которой заданный вопрос не может иметь нескольких значений“.
Принимая во внимание абсолютную функциональную значимость обстановочной афферентации для обеспечения приспособительного поведения, отметим эту афферентацию в составе схемы системной организации целостного поведенческого (пищедобывательного) акта (рис. 5).
Углубленные исследования сочетанной
роли доминирующей мотивации и обстановочной
афферентации в организации центральных
механизмов поведенческих актов показывают, что
на базе этих двух афферентаций в ЦНС формируется предпусковой
комплекс, определяющий готовность организма
развернуть не вообще некоторый поведенческий
акт, но конкретный и специфический поведенческий
акт.
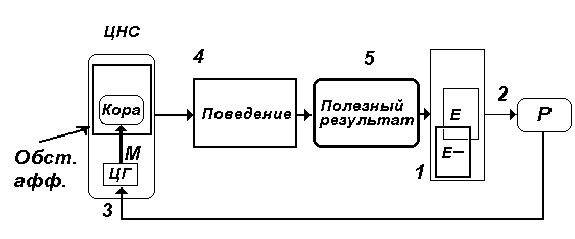
Рис. 5. Обст. афф. - обстановочная
афферентация - комплекс возбуждений от
обстановки, вместе с мотивационными
возбуждениями формирует на элементах высших
отделов ЦНС состояние предпусковой интеграции -
состояние готовности отреагировать
определенным образом. Остальное - то же, что на
рис. 4.
Несмотря на то, что физиологические предпосылки поведенческого акта готовятся до его внешнего развертывания, для экспериментатора поведенческий акт запускается как бы внезапно в строго определенный момент времени. Так, в процессе выработки пищевого условного рефлекса условнорефлекторный поведенческий акт запускается только после предъявления условного сигнала. Таким образом очевидно, что условные сигналы выступают как раздражители, вскрывающие готовность животного отреагировать на ситуацию в соответствии с уже сложившимся предпусковым комплексом. Следовательно, функциональная роль условных возбуждений может быть охарактеризована как пусковая, выводящая заранее сформированный комплекс предпусковой интеграции, созданной на базе господствующей (доминирующей) мотивации и обстановочной афферентации (рис. 6), на пороговый уровень.
Иными словами, если доминирующие
мотивационные возбуждения из состава множества
поведенческих программ мозга извлекают те
программы, которые следовало бы запустить в
первую очередь, если обстановочная афферентация
предопределяет особенности реализации
поведенческой программы применительно к
конкретной обстановке, пусковая афферентация
- это афферентация, которая активирует
сложившийся предпусковой комплекс в строго
определенный момент, наилучшим образом
отвечающий ситуации, связанной с достижением
полезного поведенческого результата.
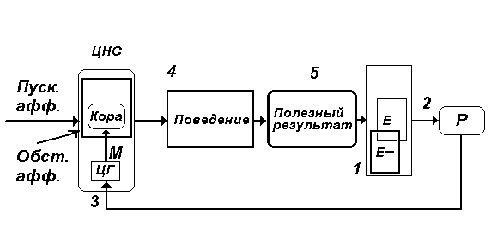
Рис. 6. Пуск. афф. - пусковая афферентация
(условный сигнал), обст. афф. - обстановочная
афферентация - комплекс возбуждений от
обстановки, вместе с мотивационными
возбуждениями формирующий на элементах высших
отделов ЦНС состояние предпусковой интеграции -
состояние готовности отреагировать строго
определенным образом. Предпусковая интеграция
вскрывается после предъявления пускового
раздражителя (условного сигнала). Остальное - то
же, что на рис. 4.
В чем заключается приспособительный смысл реагирования подопытного животного лишь на условный (пусковой) сигнал в обстановке эксперимента с выработкой условного пищевого рефлекса? Почему голодная собаки лишь по условному сигналу направляется к кормушке? Потому что обычно по условиям опыта только после включения условного сигнала животное получает доступ к кормушке. В межсигнальных промежутках доступ к кормушке заблокирован, тем самым непрерывная пищедобывательная активность становится неадаптивной, лишенной приспособительного смысла.
В лаборатории П. К. Анохина доказана особая роль лобных отделов мозга в интеграции комплекса пусковых (условнорефлекторных) афферентаций и афферентаций от обстановки. Выше отмечалось, что в межсигнальных промежутках подопытное животное с выработанным условным пищевым рефлексом обычно спокойно ведет себя, демонстрируя активность в момент включения условного сигнала. Однако, если произвести удаление лобных долей мозга у животного, характер его поведения изменяется. В этом случае у подопытного животного наблюдается непрерывная поведенческая активность, ориентированная на кормушку. Какой следует вывод? В результате лобэктомии и возникшего дисбаланса внутримозговой оценки исходного характера соотношений обстановочных и пусковых афферентаций по типу пускового раздражителя начинает действовать обстановочная афферентация.
Необходимо подчеркнуть, что мотивационные возбуждения, обстановочные афферентации, пусковые афферентации, активируя наиболее оптимальные поведенческие программы, используют аппараты памяти. Еще И. М. Сеченов отметил: "Видимое и слышимое нами всегда содержит в себе элементы уже виденные и слышанные прежде. В силу этого во время всякого нового видения и слышания к продуктам последнего присоединяется воспроизводимые из склада памяти сходственные элементы, но не в отдельности, а в тех сочетаниях, в которых они зарегистрированы в складе памяти". Весьма точное определение, подчеркивающее значение аппаратов памяти (опыта прошлого) в развитии центральных механизмов поведенческого акта.
Таким образом, необходимыый процесс начальной стадии любого поведенческого акта выступает как синтез доминирующей мотивации, обстановочной афферентации, пусковой афферентации, взаимодействующие с аппаратами памяти, сохраняющей следы прошлых значимых афферентных воздействий на мозг. Поэтому первая стадия поведенческого акта получила название стадии афферентного синтеза (рис. 7).
Нейрофизиологическую основу афферентного синтеза составляют следующие частные механизмы (по К. В. Судакову): 1. Восходящие активирующие влияния подкорковых образований на кору больших полушарий. 2. Нисходящие влияния коры на подкорковые образования. 3. Реверберация (циркуляция) возбуждений между корой и подкорковыми образованиями. 4. Механизмы конвергенции возбуждений различного сенсорного и биологического качества на нейронах ЦНС. 5. Механизмы центрального торможения.
Вторая стадия поведенческого
акта - стадия принятия решения. Как
показывает физиологический анализ, стадия
принятия решения является заключительной частью
стадии афферентного синтеза, тем не менее ее
следует рассматривать отдельно в качестве
самостоятельной стадии развертывания
функциональной системы поведенческого акта.
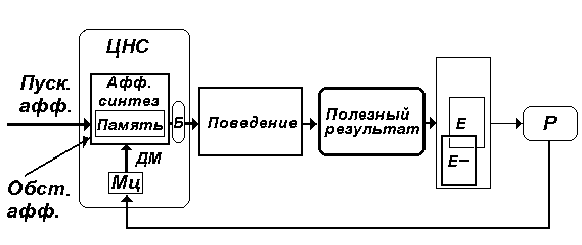
Рис. 7. Стадия афферентного синтеза -
начальная стадия любого поведенческого акта. В
ее состав входят - возбуждения доминирующей
мотивации (МЦ), обстановочная афферентация (Обст.
афф.), пусковая афферентация (Пуск. афф.),
взаимодействующие с аппаратами памяти. Б - блок
дополнительных узловых механизмов
функциональной системы центрального уровня.
Значение стадии принятия решения сводится к следующему (по П.К. Анохину):
1. “Принятие решения” освобождает организм от многочисленных степеней свободы в организации потенциальных направлений развития поведения, что обеспечивает формирование единственной совокупности эфферентных возбуждений, необходимых и имеющих приспособительный смысл для организма именно в данный момент и в данной ситуации.
2. “Принятие решения” является переходным моментом подготовительного этапа функциональной системы поведенческого акта, после которого все комбинации центральных возбуждений приобретают исполнительный, эфферентный характер.
Введем блок “принятие решения” в состав схемы функциональной системы поведенческого акта (рис. 8.).
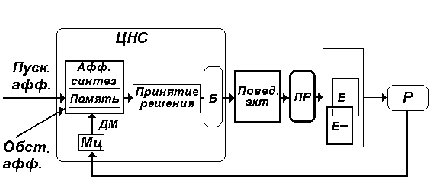
Рис. 8. Афферентный синтез и стадия
принятия решения в составе функциональной
системы поведенческого акта. Б - блок
дополнительных центральных механизмов
функциональной системы. ПР - полезный результат
Вслед за принятием решения последовательный ход событий в функциональной системе целенаправленного поведенческого акта нарушается, т. к. новый этап ее развития сопровождается параллельным развертыванием двух узловых механизмов.
С одной стороны, запускаются механизмы стадии эфферентного синтеза, с другой - формируется аппарат акцептора результатов действия (АРД) (рис. 9).
Стадия эфферентного синтеза - стадия, в
составе которой решаются вопросы сопряжения,
координации нервных, гуморальных и прочих
эфферентных механизмов управления работой
периферических эффекторов, используемых в ходе
реализации программы поведенческого акта (В. А.
Шидловский, 1982, Е. А. Юматов, Т. И. Белова, Л. С.
Ульянинский, В. И. Бадиков, 1987, К. В. Судаков, 1987, Ю.
В. Урываев, 1987). В целом, стадия эфферентного
синтеза рассматривается как динамически
развивающаяся стадия, включающая, как минимум,
этап извлечения из памяти необходимой
поведенческой программы из соответствующей
“картотеки программ” и этап реализации
избранной программы на совокупностях конечных
нейронов, имеющих выходы на периферию.
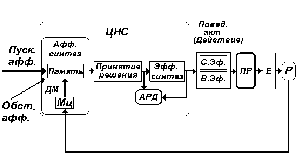
Рис. 9. Схема, иллюстрирующая развертывание в функциональной системе вслед за принятием решения двух узловых механизмов - механизмов эфферентного синтеза и акцептора результатов действия.
Выход программных возбуждений на периферические эффекторы (соматические, вегетативные) формирует внешнюю, эффекторную структуру собственно поведенческого акта, или действия, выступающего как очередной этап развития функциональной системы, продвигающий ее к достижению необходимых полезных результатов - конечной цели любого поведения (рис. 10).
Выше отмечалось, что в ходе развертывания стадии эфферентного синтеза, в составе центральных аппаратов функциональной системы параллельно идет процесс формирования акцептора результатов действия. Что это за аппарат, какова его системная роль?
Акцептор результатов действия это аппарат прогнозирования и оценки физических, химических, биологических свойств полезного приспособительного поведенческого результата путем сличения обратной афферентации от параметров реальных результатов с прогнозированной афферентной моделью этих результатов ( П. К. Анохин, 1949; А. И. Шумилина, Б. В. Журавлев, 1987).
Опыт человека убеждает в том, что
стремясь к достижению какой-либо цели, основные
параметры цели конкретизируются изначально
мысленно, т. е. идеально. В последующем идеальная
модель результатов (акцептор результатов
действия) сопоставляется с результатами
реальными. Если то, что человек “представлял”
себе совпало с тем, что получилось в
действительности - поведенческий акт
заканчивается к полному удовлетворению
субъекта, если совпадения нет, возникает чувство
досады, неудовлетворенности содеянным. Эти
эмоциональные реакции реализуются через
активацию аппарата эмоций (АЭм) (рис. 10) со стороны
акцептора результатов действия.
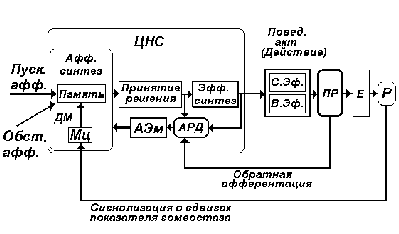
Рис. 10. Схема функциональной системы
поведенческого акта. Афферентный синтез,
принятие решения, эфферентный синтез, акцептор
действия - центральные узловые механизмы
функциональной системы поведенческого акта.
Описание в тексте.
В связи с отмеченным уместно поставить вопрос: “Подопытная собака в условнорефлекторном пищевом поведении, направляясь к кормушке, “имеет представление” о том, что в ней лежит или нет, иначе, собака “знает” или не знает, что находится
в кормушке ?”
В простом опыте можно убедиться в том, что “знает”. Для доказательства этого необходимо было оценить поведение животного в сюрпризной ситуации. В лаборатории П. К. Анохина был проведен такой опыт. По условному сигналу собака направлялась к кормушке и всякий раз получала стандартное пищевое подкрепление в виде сухарного порошка. Условный рефлекс был прочным, условная реакция у голодного животного протекала однотипно. Итак, если собака, подходя к кормушке заранее “знает”, что в ней должен быть сухарный порошок, ее поведение при внезапной подмене порошка на качественно иной корм, например, мясо должно измениться в связи с внезапно возникшей новой ситуацией. Известно, что адекватной реакцией на новизну у животных (человека) является ориентировочно-исследовательский рефлекс (рефлекс “что такое?”), поэтому задача экспериментора в данном опыте состояла в том, чтобы выяснить - у животного в момент сюрпризного соприкосновения с мясом будет ориентировочно-исследовательский рефлекс или нет?
Тщательный анализ поведения животного в рассматриваемом опыте показывает, что первый контакт животного с мясом в кормушке всегда вызывает у животного ориентировочно-исследовательский рефлекс? Поедание мяса выступает как вторичная акция, первичным оказывается ориентировочно-исследовательский рефлекс. Следовательно, подходя к кормушке собака с опережением “знает” о должном содержании подкрепления, ибо только в этом случае и может возникнуть ориентировочно-исследовательский рефлекс вследствие рассогласования между ожидаемым сухарным подкреплением и реальным мясным подкреплением при экстренной замене сухарного порошка на мясо.
Образ ожидаемого подкрепления в экспериментах с выработкой пищевых условных рефлексов, иначе центральная афферентная модель ожидаемого пищевого подкрепления, формируемая на элементах высших отделов мозга до получения реального подкрепления, - это и есть акцептор результатов действия в составе функциональной системы целенаправленного поведенческого пищевого акта. Основной процесс, который реализует акцептор результатов действия, сводится к оценке параметров полученных результатов при сопоставлении информации о результатах с предуготованной моделью этих результатов. Это происходит в ходе поступления к нейронным элементам акцептора результатов действия информации о реальных результатах поведения по каналам обратной афферентации (обратной связи) - рис. 10.
Что является аналогом акцептора результатов действия в функциональных системах гомеостатического регулирования (рис. 2)? Почему там, чаще всего, не выделяется отдельно аппарат оценки качества регулируемого процесса?
В гомеостатических функциональных системах оценка качества регулируемого процесса обеспечивается специализированными рецепторными приборами. Так, параметры осмотической константы отслеживаются осморецепторами, параметры температурной константы - терморецепторами и т. д. Функциональные свойства этих рецепторных приборов задаются генетически, т. к. должные параметры колебаний гомеостатических констант также задаются генетически. Таким образом, рецептор гомеостатической константы фактически выступает как элемент сравнения, эталон, или же акцептор результатов действия, фиксирующий отклонения константы от требуемого значения.
В поведенческих функциональных системах параметры результатов столь разнообразны, что генетический механизм создания необходимых эталонов в принципе не мог сложиться в эволюции. Вместе с тем, сложился иной - операционный механизм формирования “рецептора” результатов за счет оперативных конструкций мозга.
Наиболее сложным и дискуссионным вопросом об акцепторе результатов действия является вопрос о его нейрофизиологических механизмах. Многое здесь остается недостаточно изученным, вместе с тем общая схема развертывания этого механизма является более или менее ясной (рис. 11).
Развертывание акцептора результатов действия начинается на самых ранних этапах формирования функциональной системы, причем в этот процесс могут быть вовлечены различные по природе возбуждения, их центральные аналоги и производные, наличные и извлеченные из памяти, так или иначе отражающие вероятные свойства полезного приспособительного результата. Возможность такого подхода представляет концепция П. К. Анохина о биологических корнях приспособительных поведенческих актов, в частности, тезис о примате сигнальности как наиболее характерном и адаптивном признаке всякого целостного поведения - "каждый ... поток импульсов может стать сигнальным раздражителем для последующего, если только "последующий" составляет существенную сторону в жизни организма" ( П. К. Анохин, 1968).
Обратимся к наиболее ранней стадии
функциональной системы - к стадии афферентного
синтеза, основой которой выступает доминирующая
мотивация. По результатам исследований,
проведенных К. В. Судаковым (1978, 1979), Ю. А. Макаренко
(1980) было показано, что мотивационные
возбуждения, находясь в цепи стабильной
последовательности событий (потребность -
мотивационные возбуждения - поведение,
направленное на удовлетворение мотивации, -
полезный результат) при многократном повторении
этих событий начинают выступать сигналом
будущего подкрепления. Следовательно,
мотивационные возбуждения на самых ранних
стадиях поведенческого акта уже закладывают
основы формирования центрального комплекса
возбуждений, предвосхищающего параметры
будущего полезного поведенческого результата, т.
е. акцептора результатов действия.
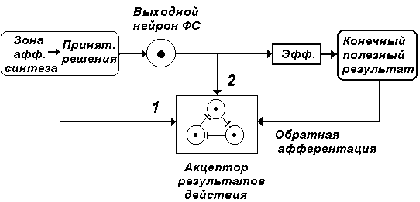
Рис. 11. Схема, иллюстрирующая механизм
формирования акцептора результатов действия
(АРД). 1.Возбуждения, отражающие участие
процессов, производных от возбуждений
(мотивационных, обстановочных, пусковых) стадии
афферентного синтеза. 2. Коллатеральные
возбуждения выходных нейронов функциональной
системы, выступающие как опережающий
информационный эквивалент конечных полезных
результатов.
Дальнейшее развитие функциональной системы сопровождается поступлением в мозг возбуждений иного качества. Это обстановочные возбуждения, пусковые возбуждения. Несомненно, мотивационные возбуждения, активируя аппараты долговременной памяти, извлекают многое, что ранее связывалось с удовлетворением возникшей мотивации, формируя первичный комплекс акцептора результатов действия. Однако, обстановочная афферентация, пусковые раздражители - ближайшие предвестники подкрепления, воздействуя на мозг, извлекают из памяти новую дополнительную информацию, обогащающую акцептор результатов действия, что делает образ ожидаемых результатов более осязаемым в силу перехода организма в состояние, когда вероятность успешной реализации центральных поведенческих программ и достижение полезных результатов повышается почти до единицы.
Таким образом, можно заключить, что становление акцептора результатов действия на начальных стадиях развертывания функциональной системы связано со стадией афферентного синтеза - рис. 11, позиция 1.
Совокупность стартовых внутримозговых процессов функциональной системы, влияющих на формирование акцептора результатов действия, может быть большей или меньшей, однако, новое качество этим процессам придает выход командных возбуждений на двигательные эффекторы.
Прежде всего подчеркнем, что совокупный эфферентный разряд выходных нейронов функциональной системы, посылающих команду на периферию, информационно всегда отражает содержание реализуемой поведенческой программы и заключает в себе импульсный информационный эквивалент того, что должно произойти на эффекторах функциональной системы. В функциональной системе все ориентировано на достижение поставленной цели. Отсюда - совокупность центральных процессов представляет собой центральный информационный эквивалент будущих результатов действия, совокупность импульсаций от рецепторых приборов, передающих в центральную нервную систему возбуждения об отдельных параметрах полученных результатов, есть афферентный информационный эквивалент полученных результатов, совокупный эфферентный разряд на периферию, призванный обеспечить достижению конечных полезных результатов, являет собой эфферентный информационный эквивалент полезных результатов. На основании изложенного П. К. Анохин (1968) выдвинул гипотезу о том, что в качестве заключительной составляющей общего процесса формирования акцептора результатов действия выступают коллатеральные возбуждения выходных нейронов функциональной системы, копирующих структуру выходного командного потока, но остающихся в распоряжении центральных аппаратов функциональной системы (рис. 11, позиция 2).
По коллатералям выходных моторных нейронов функциональной системы копии эфферентных команд, фактически копии программы поведенческого акта, распространяются на нейронные комплексы мозга - на мультисенсорный вставочный аппарат моторной коры, на нейроны ассоциативных полей коры, подкорковых ганглиев, ретикулярной формации, мозжечка и т. д., где в последующем происходит встреча и взаимодействие этих возбуждений с возбуждениями афферентной природы, потенциально отражающих содержание полученных результатов. Возможность таких соотношений между эфферентными коллатеральными и афферентными периферическими возбуждениями доказана в конкретных нейрофизиологических опытах (рис. 12).
Очередным этапом в развертывании функциональной системы вслед за “действием” является получение полезных результатов и оценка достигнутых результатов на элементах акцептора результатов действия.
Можно выделить два крайних варианта оценки результатов того или иного поведенческого акта.
1. Поведенческий акт завершается успешно, обратная информация о полученном результате поступает на элементы акцептора результатов действия, при этом ожидаемая модель результатов совпадает с тем, что было получено в ходе поведенческого акта. В этом случае акцептор результатов действия активирует аппарат биологически положительных эмоций, что приводит к формирования эмоции удовлетворения свершенным. Положительная эмоция в данной ситуации выступает как сигнал успешного завершения поведенческого акта, поведенческий акт на этом заканчивается.
2. Поведенческий акт завершается с ошибками в достижении необходимого результата, при этом ожидаемая модель результатов не совпадает с тем, что было получено в ходе поведенческого акта - на элементах акцептора результата действия возникает реакция рассогласования, при этом активируется аппарат биологически отрицательных эмоций. Степень рассогласования может быть различной. В зависимости от этого внешние проявления неудачного завершения поведенческого акта могут варьировать от ориентировочно-исследовательской реакции до состояния биологически отрицательной реакции с выраженным негативным эмоциональным компонентом. Адаптивный смысл включения аппаратов отрицательных эмоций в случае неполучения необходимых полезных результатов заключается в том, что биологически отрицательная реакция выступает как стимул, организующий повторный, более совершенный поведенческий акт.
Следует обратить внимание на чисто
медицинские аспекты биологически отрицательных
реакций человека (реакций неудовлетворения, или
неудовольствия), отражающих состояние конфликта
при длительном или невозможном достижении
потребного результата. В конфликтной ситуации
при усилении отрицательного эмоционального
напряжения последнее может переходить в
состояние эмоционального стресса,
который становится основой возникновения и
развития нарушений в работе целого

Рис. 12. Конвергенция коллатеральных эфферентных (командных) возбуждений моторных нейронов пирамидного тракта и афферентных возбуждений (возбуждений в составе обратной афферентации) на корковом нейроне. Сверху вниз - реакции нейрона коры головного мозга на раздражение коллатералей аксонов пирамидного тракта (ПТ), на вспышку света, на соматическое воздействие (боль), вестибулярное раздражение.
организма в форме артериальной гипертензии патологических изменений деятельности сердца, иммунодефицитных состояний, язвенных поражений желудочно-кишечного тракта (К. В. Судаков).
Необходимо подчеркнуть, что качество поведенческих результатов окончательно проверяется по критерию восстановления параметров той или иной гомеостатической константы, сдвиг которой создал первичные предпосылки поведенческого акта. Применительно к пищедобывательному поведенческому акту качество принятой пищи лишь оперативно оценивается на элементах акцептора действия, решающая оценка достигнутого результата производится по параметрам изменения концентрации питательных веществ в крови. Можно считать, что принятая пища удовлетворяет биологическим критериям, если после приема пищи имеет место нормализация концентрации питательных веществ в циркулирующей крови и последующее адекватное включение питательных веществ в процессы обмена веществ в организме.
Несмотря на то, что рассмотренная системная архитектура физиологически была спроецирована на условнорефлекторный пищедобывательный акт, предложенная схема является универсальной, в одинаковой мере приложимой для описания операционной структуры поведенческих актов любой биологической или социальной модальности (П. К. Анохин, К. В. Судаков).
В заключение настоящего раздела приводим краткое описание общей последовательности развертывания узловых механизмов функциональной системы целенаправленного поведенческого акта (рис. 13).
1. Стадия афферентного синтеза.
2. Стадия принятия решения.
3а. Стадия эфферентного синтеза.
3б. Стадия формирования акцептора результатов действия.
4. Стадия выхода программных импульсаций на эффекторы - стадия действия (собственно поведенческий акт).
5. Стадия получения полезных результатов, формирование потока обратной афферентации.
6. Стадия оценки параметров полученных
результатов на элементах акцептора результатов
действия (сопоставление афферентной модели
полезных результатов с реально полученными в
поведенческом акте полезными результатами).
Запуск аппарата положительных эмоций в ситуации совпадения
акцептора результатов действия с обратной
афферентацией. Завершение развития
функциональной системы при адекватном
воздействии поведенческого результата на
параметры гомеостатической константы. В
ситуации несовпадения акцептора
результатов действия с обратной афферентацией -
запуск ориентировочно-исследовательского
рефлекса или биологически отрицательной реакции
с последующим формированием новой
функциональной системы более адаптивного
поведенческого акта.
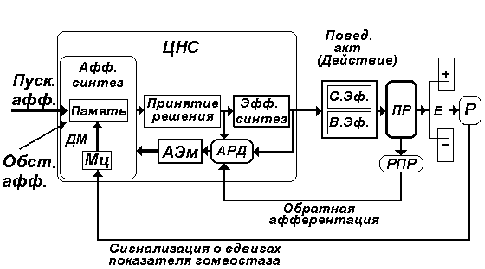
Рис. 13. Блок-схема поведенческого акта с позиций теории функциональной системы П. К. Анохина. В сравнении со схемой рис. 12 введен блок РПР - совокупность рецепторов, оценивающих параметры полезного результата, достигнутого в рамках целенаправленного поведенческого акта.
ЛИТЕРАТУРА
Анохин П.К. Внутренее торможение как проблема физиологии. -М.:Медгиз, 1956. -378с.
Анохин П. К. Эмоции // БМЭ, 2 издание.
Анохин П. К. Предисловие к книге Э. Гельгорна, Дж. Луфборроу “Эмоции и эмоциональные расстройства”- М.: Мир, 1966.- С. 5-18.
Анохин П. К. Биология и нейрофизиология условного рефлекса. - М.: Медицина, 1968.- 540 с.
Анохин П. К. Очерки по физиологии функциональных систем. - М.: Медицина, 1974. - 446 с.
Макаренко Ю. А. Системная организация эмоционального поведения. - М.: Медицина,1980. - 157 с.
Павлов И. П. Двадцатилетний опыт изучения высшей нервной деятельности (поведения) животных. Условные рефлексы. Изд. 2, доп.- Л.:Госизд., 1924.
Павлов И. П. Полное собрание сочинений. Объективное изучение высшей нервной деятельности животных. Полн. собр. соч. Изд.2.-Т. 3, кн. 2.- М. -Л.: АН СССР, 1951.- 433 с.
Судаков К. В. Биологические мотивации. - М.: Медицина,1971. - 303 с.
Судаков К. В. Общая теория функциональных систем. - М.: Медицина, 1984.-224 с.
Судаков К. В. Функциональные системы организма. Под ред. К. В. Судакова. -М.: Медицина, 1987.- 432 с.
Судаков К. В. Избранные лекции по нормальной физиологии.-М.:Эрус,1992.-243 с.
Урываев Ю. В. Высшие функции мозга и поведение человека. -М. - 1996. - 200 с.
Кафедра нормальной физиологии
Смоленская государственная меддицинская академия
Поступила в редакцию 13.01.98.