Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 11. -
Вып. 3. - 2012. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/TITL-35.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/cont.htm
УДК
572. 1/.4
Африка – Европа: анализ дивергенции
краниометрических признаков
Ó
2012 г. Харитонов В. М.
В работе продолжен анализ пространственной
структуры краниометрических признаков гоминид, охватывающую три части света
(Африку, Азию, Европу). Результаты получены с помощью канонического анализа,
проведенного на основе компьютерной системы HomoBase-Kanoklass, созданной Е. В.
Дерябиным. Материалом для работы послужили данные по краниометрии ископаемых
гоминид плейстоцена Африки, Азии Европы, оформленные в виде РС базы данных,
включающих измерения черепа 250 гоминид. Обсуждаются результаты анализа 82
ископаемых гоминид трех частей света (австралопитеки, H. habilis, H.
rudolfensis, H.ergaster, H. erecrus, H. heidelbergensis, H. neanderthalensis,
архаичный H. sapiens). Канонический анализ объективно отражает сходство и
различие ансамблей краниометрических признаков ископаемых гоминид. Конкретно
исследовались особенности дивергенции (трансгрессии) краниометрических
признаков географических кластеров плейстоценовых гоминид Африки и Европы. В
нашей классификации они проявляется в вариантах географических группировок
Африка+Европа; Азия - отдельно (11,31%) и Африка+Азия+Европа (29, 92%). Суммарная
величина двух вариантов – 41,23%. т.е. меньше половины от всех изученных
случаев. Встречаемость случаев трансгрессии краниометрических признаков у
гоминид Африки и Европы меньше, чем у Африки и Азии, что соответствует ожиданию
(Европа была заселена мигрантами из Африки позже чем Азия по данным
палеонтологии гоминид). Трансгрессия краниометрических признаков ископаемых гоминид
Африки и Европы выглядит следующим образом: 1. Африка+Европа; Азия отдельно. Результат
получен при анализе: мозговой отдел
– неандертальцы (гейдельбержцы),
кальвариум – австралопитеки, лицевой отдел – неандертальцы, нижняя челюсть –
эректусы. 2. Африка+Европа+Азия. Результат получен при анализе: мозговой отдел
- неандертальцы, гейдельбержцы,
кальвариум – австралопитеки, лицевой отдел – эректусы, гейдельбержцы, нижняя
челюсть – эректусы, неандертальцы. Чаще всего, ископаемых гоминид Африки и
Европы объединяют анализы: неандертальцев, эректусов, гейдельбержцев,
австралопитеков. Вариант пространственного расположения географических
группировок ископаемых гоминид Африка+Европа; Азия отдельно в изученной
совокупности канонических анализов встречается редко. Он выявлен при изучении
гоминид нижнего, среднего и верхнего
плейстоцена. Можно лишь предположить, что трансгрессия признаков мозгового
отдела черепа у гоминид Африки и Европы выражена в большем числе случаев чем у
отделов висцерального черепа (собственно лицевой отдел черепа, нижняя челюсть).
Вариант Африка+Европа+Азия – встречается в совокупности графиков ископаемых
гоминид, полученных с помощью канонического анализа, в большем числе случаев
чем вариант – Африка+Европа , Азия отдельно. По частоте проявления трансгрессии
признаков черепа гоминид Африки и Европы отделы черепа расположены так:
мозговой отдел, нижняя челюсть → лицевой отдел → кальвариум.
Трансгрессия признаков по частоте проявления у гоминид Африки и Европы
встречается чаще в следующих случаях: мозговой отдел (нижний – верхний
плейстоцен), нижняя челюсть (нижний- верхний плейстоцен) → лицевой отдел
(нижний- верхний плейстоцен) → кальвариум (эоплейстоцен-нижний плейстоцен
). По результатам проведенной работы филогенетические связи африканских и
азиатских гоминид представляются более древними, чем африканских и европейских.
Ключевые слова: антропология
(физическая антропология), палеоантропология, гоминиды Африки, гоминиды Европы,
дивергенция признаков
Эволюционные взаимоотношения
ископаемых гоминид Африканского материка и Европейского континента являются
одной из актуальных проблем палеоантропологии палеолита. В науке обсуждаются
вопросы: а) древности и эволюционной природы первых насельников Европы, б)
происхождения европейских неандертальцев, в) возможного происхождения сапиенса
в Европе и т.д.
Приведем литературную
справку о палеоантропологии Европы. В период 1-1,5 млн. лет назад архантропы
заселили южные районы Европы и Азии, вплоть до Индонезии; в Австралию и Америку
они не мигрировали [Дробышевский, 2009].
А. П. Деревянко менее
категоричен. Он пишет об эректусе как виде, который не только широко расселился
в Африке, Азии и, вероятно, Европе, но и существовал около 1,5 млн лет. В
Европе, продолжает он, нет остатков гоминид такой же древности, как в Грузии,
Восточной и Юго-Восточной Азии. Дискуссионными являются древность и рукотворность
артефактов в европейских местонахождениях Валлоне, Баранко-Леон, Валлоне.
А. А. Зубов и С. В. Васильев
[2006] относят время заселения Европы к рубежу 800-700 тыс. лет. А.П.Деревянко
[2009] на основе археологических свидетельств допускает возможность заселения
Европы более 1 млн. лет назад. Вопрос заключается в том, какова видовая
принадлежность первых европейских насельников? А.А.Зубов и С.В.Васильев [2006]
относят их к ранним формам гейдельбергского человека, а не к эректусам.
Древнейшие гоминиды Европы
открыты в местонахождении Атапуэрке (Испания), они размещаются в диапазоне
1,2-0,5 млн. лет. Ископаемые останки гоминид, открытые в Гран-Долине,
древностью в 800 тыс.лет, позволили предположить существование на юге Европы
нового вида H.antecessor. Авторы этого предположения
считают, что он имеет африканские корни, связан эволюционными корнями с
эргасторами, и является предком и неандертальцев Европы, и сапиенса. Таким
образом, гейдельбержцы Африки и Европы выводятся из родословной сапиенса.
Первые представители H. antecessor в среднем плейстоцене
мигрировали в Европу и дали начало европейским неандертальцам. Африканские
представители этого вида стали предковыми для промежуточных видов – предков H.sapiens.
В соответствии с другой точкой зрения H. аntecessor мог быть потомком дманисцев,
если допустить возможность видообразования в Европе. Мы помним об африканских
корнях последних и отнесении их к архантропам. Мигранты второй волны – архантропы
могли вытеснить или ассимировать H. аntecessor. Наконец, есть еще одна
точка зрения: H. аntecessor является результатом
кладогенеза, произошедшего после
ашельской миграционной волны [Деревянко,2009].
Указанный автор пишет, что
имеющиеся на сегодняшний день археологические и антропологические материалы убеждают
в том, что миграций из Африки в Евразию и обратно в раннем палеолите было
немного. Можно проследить только два глобальных миграционных процесса. Каждая
такая миграция детерминировалась глубокими причинами. По А. П. Деревянко
[2009], попытки объяснить возникающие трудноразрешимые проблемы, в т.ч.
некоторые филогенетические, древнейшими миграционными процессами на сегодняшнем
уровне знаний нельзя считать продуктивными. Расселение человека в Европе началось
в то же время, что и в Азии. Это был единый процесс миграции из колыбели
человечества - Африки. Но архантропы быстрее всего заселяли близкие им по
экологическим условиям территории южной части Азиатского континента и в течение
сравнительно короткого времени вышли к Тихому океану. Значительно медленнее они
могли продвигаться на север в Европу через Ближний Восток, Малую Азию, Балканы
и т.д. или же через Северный Кавказ. Наиболее короткий путь человека из Африки
в Европу мог быть по сухопутному мосту. Но когда Европа соединилась с Африкой
или, по крайней мере, существовали мелководные, т.е. преодолимые для
архантропа, проливы при понижении уровня Мирового океана - эти проблемы требуют
своего разрешения.
На современном уровне знания
самым древним (бесспорно) палеолитическим местонахождением в Западной Европе является
Сима-дель-Элефанте (Испания), но это, конечно, не исключает открытия новых,
более древних стоянок или подтверждения древности некоторых дискуссионных. Кто
были первые пришельцы в Европу? Этот вопрос остается дискуссионным. Очень
вероятно, что в Западную и Центральную Европу первыми проникли Homo georgicus
[Деревянко,2009].
А. П. Деревянко [2009]
обращает внимание на то, что многие антропологи отмечают значительную
эректоидную примесь у древнего населения Европы. Об этом свидетельствует черепная
коробка из Чепрано (Италия). Поэтому отрицание роли Homo erectus в эволюционном
процессе в Европе неоправданно.
Вторая глобальная миграция
человека из Африки (с Ближнего Востока?) в Европу произошла около 600 тыс.
л.н. Очень вероятно, что вторая миграционная
волна из Африки в Европу была связана с представителями Homo heidelbergensis,
для которых характерно сочетание неандерталоидных и сапиентных черт.
В связи со второй
миграционной волной встает вопрос о судьбе автохтонного населения Европы. На одних
местонахождениях среднего плейстоцена преобладает ашельская индустрия, на
других прослеживается бифасиальная и галечная, на третьих отсутствуют рубила.
Видимо, с приходом новой миграционной волны в Европе происходил процесс не
замещения, а аккультурации. В этом отношении очень важны находки на
местонахождении Сима-де-лос-Хуэзос (Испания) древностью более 530 тыс. лет.
Антропологические исследования около 4 тыс. человеческих костей показали, что
эти архантропы существенно отличались от тех, чьи останки найдены в горизонте
ТД6 Гран-Долины, и были близки к неандертальцам. Палеоантропологические
материалы местонахождения Сима-де-лос-Хуэзос доказывают, что Европа была очагом
местной эволюционирующей ветви, впоследствии давшей начало « классическому»
неандертальцу. Многие вопросы, связанные с двумя волнами миграции человека из
Африки в Европу, остаются дискуссионными [Деревянко,2009].
С. В. Дробышевский [2009]
отмечает, что существует факт явной неоднородности уровня прогресса синхронных
гоминид в разных регионах. Так, в Юго-Восточной Азии, особенно в Индонезии, в
период 130-400 тыс. лет назад часты находки, определяемые как Homo erectus; в
Китае есть много находок, определяемых
как "архаический Homo sapiens", хотя их признаки не так сапиентны, как у синхронных им
африканских гоминид. В Европе местами обитали довольно архаичные люди, но
преобладали, видимо, Homo heidelbergensis (эти
формы определяются как
"атипичные неандертальцы" или "пресапиенсы").
Отличие неандертальцев от
современных людей были весьма существенны. Они тем более значительны, что на
протяжении не менее 5 тысяч лет неандертальцы сосуществовали в Европе с людьми
современного вида. Были ли они нашими предками? Сейчас нет однозначного ответа
на вопрос о судьбе неандертальцев, на этот вопрос учёные отвечают по-разному,
есть три основные версии.
С момента около 40-45 тысяч
лет назад люди совершенно современного облика, только несколько более
массивные, чем мы, – неоантропы – известны практически со всей территории
ойкумены – из Африки, Европы, Азии и Австралии.
Многие биологические
особенности древнейших верхнепалеолитических людей позволяют предположить, что
они пришли в Европу из тропических областей. Длинные конечности, высокий рост,
вытянутые пропорции тела, крупные челюсти, вытянутая мозговая коробка схожи у
современных тропических популяций и кроманьонцев. Последние отличаются только
большими размерами костей, сильным рельефом черепа, более грубыми чертами. По
наиболее обоснованной сейчас версии, современный вид человека сформировался в
Африке между 200-160-100 и 45 тысячами лет назад. Между 80 и 45 тысячами лет
назад ограниченное число людей вышло из Восточной Африки в районе
Баб-эль-Мандебского пролива или, менее вероятно, Суэцкого перешейка. Они стали
расселяться сначала по южным берегам Евразии – вплоть до Австралии, – а после
на север, в области, заселённые неандертальцами, о возможной судьбе которых
сказано выше.
В одной из прошлых работ
[Дерябин, Харитонов, 2003] мы касались взаимоотношений конкретных ископаемых
гоминид Африки и Европы, которые маркируют «афро-европейский» этап в
происхождении сапиенса по «афро-европейской гипотезе». Речь идет о миграциях
предшественников неандертальцев из Африки в Европу [Brauer,1984]. Опорными находками для предположения об
«афро-европейском» этапе в миграциях гоминид Африки являются представители H.heidelbergensis (гоминиды Брокен-Хилл в
Африке, Араго, Петралона – в Европе). Признаки нейрокраниума позволяют видеть,
что Брокен-Хилл безоговорочно не ближе к Петралоне чем к архантропам, Салданье,
Штейнгейму или поздним западноевропеским неандертальцам. При этом гейдельбержец
Брокен-Хилл среди европейских гоминид расположен ближе к поздним
неандертальцам, чем к ранним и кроманьонцам. При этом по признакам нейрокраниума гейдельбержцы Брокен-Хилл и Салданья одинаково
отличны от кроманьонцев.
Анализ объединенного
ансамбля признаков мозгового и лицевого отделов показал, что KNM-3733 и
Брокен-Хилл (Родезия)ближе расположены к представителям H.neanderthalensis, чем
Бодо . Таким образом, единичные наблюдения позволяют осторожно предположить,
что H.ergaster достаточно близок к верхнеплейстоценовым гоминидам, но при этом
дальше чем большинство H.heidelbergensis.
Европейская форма т.н.
"пренеандертальца" Араго (Франция) (0,4 млн.лет), относимая к
H.heidelbergensis, по признакам нижней челюсти близка к H.erectus Атлантропу
III(Алжир). Араго демонстирует большую близость к некоторым неандертальцам чем
представители H.erectus.
Вид H. heidelbergensis
включает в себя и внеевропейские формы того же эволюционного уровня. Брокен-Хилл
(0,3 млн. лет) в Африке относится к их числу. Проведенный анализ выявил
тяготение гейдельбержца Брокен-Хилл к неандертальцам, но и достаточное
различие между ними . По признакам нейрокраниума родезиец отличен от более
поздних гоминид явно, а по признакам лицевого отдела проявляет отличие от кроманьонцев
аналогичное неандертальцам. В недалеком прошлом родезиец часто
классифицировался как вариант "внеевропейских палеоантропов", в
определенной степени сходный с Нгандонг (Ява) [Рогинский, 1966]. Наш результат
более близок к данному результату, чем к отнесению Брокен-Хилл к
представителям "переходного протосапиентного" пласта гоминид [Булыгина,
Хрисанфова, 1998]. Хотя авторы отмечают черты сходства Брокен-Хилл с классическими
европейскими и переднеазиатскими вариантами палеоантропов.
Гоминид Салданья (Родезия)
не тождественна Брокен-Хилл по признакам нейрокраниума, данный гоминид
располагается между представителями неандертальского вида, H.erectus и
H.heidelbergensis. Можно наблюдать тяготение Брокен-Хилл и Салданьи к
H.erectus. Салданья относилась ранее также к категории "внеевропейских
палеоантропов" [Рогинский, 1966].
По признакам лицевого отдела
черепа Брокен-Хилл ближе к Петралоне чем к неандертальцам (кроме архаичного
сапиенса Схул) и кроманьонцам. Среди схульцев гейдельбержцы Брокен-Хилл и
Петралона ближе к Схул IY и по-разному расположены по отношению к архаичному
сапиенсу (?) Джебель-Ирхуд I.
Признаки нижней челюсти
выявляют относительную близость европейских гейдельбержцев Араго XIII и Араго
III ( 0,4 млн лет) к H.erectus Атлантропу III (0,7 млн.лет) и европейским
неандертальцам, позволяя отнести их к группе "пренеандертальцев"
Европы. Таким образом, проведенные расчеты с помощью канонического анализа
cоотносимы с обсуждаемой моделью африканской прародины сапиенса, но не по всем
категориям признаков черепа.
Материалы и
методы исследования
В данной работе продолжен
анализ пространственной структуры краниометриче-ских признаков гоминид,
охватывающую три части света (Африку, Азию, Европу) с помощью результатов
канонического анализа, проведенного на основе системы HomoBase-Kanoklass,
созданной профессором Е.В.Дерябина. Канонический анализ объективно отра-жает
сходство и различие ансамблей краниометрических признаков ископаемых гоминид.
Материалом для работы
послужили данные по краниометрии ископаемых гоминид плей-стоцена Африки, Азии
Европы, оформленные в виде РС базы данных, разработанной д.б.н. В.Е.Дерябиным.
Указанная база включает измерения черепа 250 гоминид. В на-стоящей работе
обсуждены результаты анализа 82 ископаемых гоминид трех частей света. Имеются в
виду: а) австралопитеки – 3 вида; б) H.habilis-1, в) H. rudolfensis-1, г)
H.ergaster- 1, д) H.erecrus-29, е) H.heidelbergensis-12, ж)
H.neanderthalensis-23, з)архаичный H.sapiens- 12.
Результаты и
обсуждение
1. Здесь мы обсудим вопрос особенностей дивергенции (трансгрессии)
краниометрических признаков географических группировок ископаемых гоминид
Африки и Европы, выявленные в результате канонического анализа. В нашей классификации они проявляется в
вариантах «г» и «б», т.е. в комбинациях географических группировок
Африка+Европа; Азия - отдельно (11,31%) и Африка+Азия+Европа (29, 92%).
Суммарная величина – 41,23%. т.е. меньше половины от всех изученных случаев.
Встречаемость случаев трансгрессии краниометрических признаков у гоминид Африки
и Европы меньше, чем у Африки и Азии, что соответствует ожиданию (Европа была
заселена мигрантами из Африки позже). Рассмотрим варианты взаимного
пространственного расположения географических групп гоминид.
Вариант «г». Африка+Европа;
Азия отдельно.
Явления трансгрессии признаков черепа гоминид было обусловлено
следующими отделами черепа (МО=2; МО+ЛО=0; ЛО=1; НЧ=1).
Для признаков мозгового
отдела явление трансгрессии выявлено при анализе представителей видов
гейдельбергского человека и неандертальского человека, т.е. гоминид среднего и
верхнего плейстоцена. Резервом для данного варианта взаимного положения гоминид
разных континентов являются много численные случаи? + Европа; Азия отдельно.
Для мозгового отдела их около 29. Этот вариант выявлен при анализе
питекантропов (17), неандертальцев (9) и гейдельбержцев (3), т.е. ископаемых
гоминид нижнего, среднего и верхнего плейстоцена (рис.1).
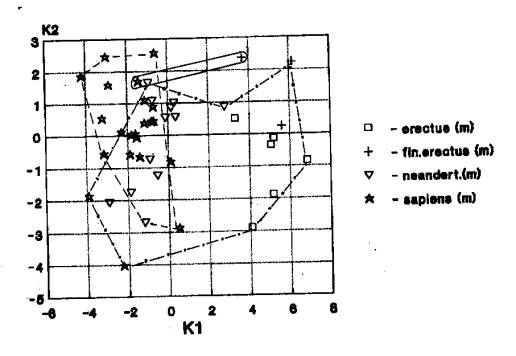
На рис.1 мы видим частичное пространственное
совмещения значений краниометрических признаков мозгового отдела гоминид Африки
и Европы.
Для признаков мозгового и
лицевого отделов черепа вместе изучаемый вариант взаимного положения
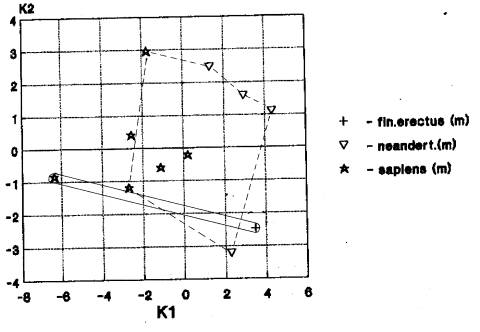
географических групп гоминид не наблюдается , лицевого отдела – при анализе неандертальцев (рис.2), нижней
челюсти – при анализе питекантропов. На рис.2 мы видим трансгрессию
краниометрических признаков (лицевой отдел) у гоминид Африки и Европы.
Обобщим полученные
результаты. Вариант пространственного расположения географических группировок
ископаемых гоминид Африка+Европа; Азия отдельно в изученной совокупности
канонических анализов встречается редко. Он выявлен при изучении гоминид
нижнего, среднего и верхнего плейстоцена. Можно лишь предположить, что трансгрессия
признаков мозгового отдела черепа у гоминид Африки и Европы выражена в большем
числе случаев чем у отделов висцерального черепа (собственно лицевой отдел
черепа, нижняя челюсть).
Вариант «б». Африка+Европа +
Азия.
Явления трансгрессии признаков черепа гоминид было обусловлено
следующими отделами черепа (МО=23; МО+ЛО=4; ЛО=10; НЧ=23).
Мы видим, что явление
трансгрессии краниометрических признаков у мозгового отдела гоминид Африки и
Европы выражено сильно. Оно выявлено при анализе неандертальцев (8),
гейдельбержцев (6), эректусов (5), архаичных сапиенсов (3) и ранних Номо (1).
Также достаточно часто встречен вариант трансгрессии? + Европа + Азия (13),
который может послужить резервом для предедущего. Он выделен при анализе
архаичных сапиенсов (5), неандертальцев (6), гейдельбержцев (3). Указанные два
варианта трансгрессии присущи гоминидам в широком временном интервале – от
нижнего до верхнего плейстоцена (рис.3).
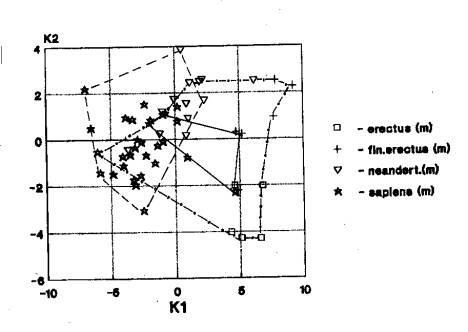
На рис.3 мы видим
пространственное совмещение областей значений краниометрических признаков
(мозговой отдел) гоминид Африки, Европы и Азии (41).
Кальвариум. Наблюдался один
вариант - трансгрессия признаков черепа гоминид трех частей света (4). Он
проявился при анализе австралопитековых и эректуса. Допускаем, что изменчивость
признаков черепа у австралопитековых столь велика, что включает в себя
внутривидовую изменчивость ископаемых гоминид трех частей света (рис.4).
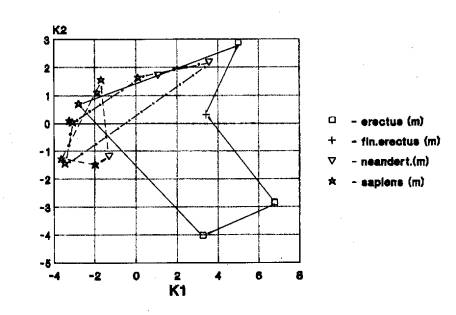
На рис.4 мы видим
трансгрессию краниометрических признаков мозгового и лицевого отделов у гоминид
Африки, Европы и Азии.
Лицевой отдел черепа. Наблюдается вариант
трансгрессии признаков черепа у ископаемых гоминид Африки и Европы (10). Он выявлен при анализе эректусов (3),
гейдельбержцев (3), неандертальцев (2) и архаичного сапиенса (1) (рис.5).
Второй вариант - ?+Азия+Европа (3).
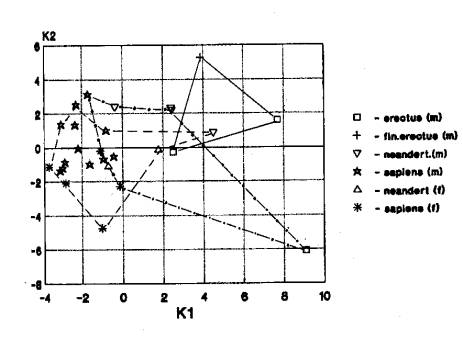
На рис.5 мы видим
трансгрессию краниометрических признаков лицевого отдела гоминид Африки, Европы
и Азии.
Нижняя челюсть. Трансгрессия краниометрических
признаков гоминид Африки и Европы выражена значительно (23). Она выявлена при анализе
эректусов (13). неандертальцев (70), гейдельбержцев (3). Спорный вариант -
?+Азия+Европа выявляется при анализе неандертальцев (1) (рис.6).
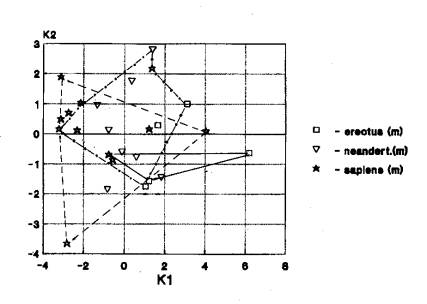
На рис. 6 мы видим
трансгрессию краниометрических признаков нижней челюсти у гоминид Африки,
Европы и Азии.
Обобщим полученные
результаты. Вариант – Африка+Европа+Азия – встречается в совокупности графиков
ископаемых гоминид, полученных с помощью канонического анализа, чаще чем
вариант – Африка+Европа , Азия отдельно. В обсуждаемом варианте трансгрессию краниометрических
признаков различные отделы черепа демонстрируют в следующем соотношении:
мозговой отдел (МО) - 23 случая, кальвариум (МО+ЛО) – 4 случая, лицевой отдел
(ЛО)-10 случаев, нижняя челюсть (НЧ) – 23 случая. Таким образом, по частоте
проявления трансгрессии признаков черепа гоминид Африки и Европы отделы черепа
расположены так: мозговой отдел, нижняя челюсть → лицевой отдел →
кальвариум. Трансгрессия признаков по частоте проявления у гоминид Африки и
Европы встречается чаще: мозговой отдел (нижний – верхний плейстоцен), нижняя
челюсть (нижний-верхний плейстоцен) → лицевой отдел (нижний- верхний
плейстоцен) → кальвариум ( нижний плейстоцен - эоплейстоцен).
Актуальным является вопрос,
как полученные с помощью канонического анализа данные согласуются с имеющимися
в палеоантропологии и археологии касательно очередности заселения африканскими
мигрантами Азии и Европы.
2. Межрегиональные связи
стадиальных групп ископаемых гоминид Африки и Азии.
Эволюция гоминид
прослеживается между конкретными стадиальными и внутристадиальными группами
ископаемых гоминид. Соотношение верхнеплейстоценовых гоминид Африки и Европы
особенно интересно, так как маркирует возможные филогенетические связи
ископаемых гоминид Африки и Европы, а также показывает на генезис предшественников
неандертальцев и сапиенса (?), совпадающий в географическом аспекте с миграциями
гоминид из Африканского материка в Европу.
Анализ 191 графика
результатов канонического анализа краниометрических признаков ископаемых
гоминид Африки и Европы продемонстрировал следующие варианты морфологической
близости.
1.
Выявлено
тяготение гейдельбержцев Африки и неандертальцев Европы.
2.
Броккен-Хилл
– Феррасси, Шапель-о-Сен – череп без нижней челюсти (5 случаев); Броккен-Хилл,
Бодо – Шапель-о-Сен, Саккопасторе,
Монте-Чирчео – лицевой отдел (4 случая); Броккен-Хилл – Феррасси,
Шапель-о-Сен, Мустье – мозговой отдел (5 случаев).
3.
Выявлено
тяготение гейдельбержцев Африки и Европы. Броккен-Хилл – Петралона, Фонтешевад-
лицевой отдел (4 случая); Выявлено тяготение эргастеров Африки и неандертальцев
Европы. КНМ 3733 – Шапель-о-Сен- череп без нижней челюсти (1 случай); КНМ 3733-
Саккопасторе – мозговой отдел (1 случай); КНМ 3733- Гибралтар 1 – лицевой отдел
(2 случая).
4.
Выявлено
тяготение питекантропов Африки и неандертальцев Европы. Атлантроп 2
a.
Монтморен
– нижняя челюсть (2 случая).
5.
Выявлено
тяготение питекантропов Африки и гейдельбержцев Европы. Атлантроп 1 и Атлантроп
3 – Араго 3 и Мауэр – нижняя челюсть (2 случая).
6.
Выявлено
тяготение питекантропов Африки и кроманьонцев Европы. Атлантроп 2 – Оберкассель
– нижняя челюсть (1 случай).
7.
Выявлено
тяготение неандертальцев Африки и гейдельбержцев и неандертальцев Европы.
Джебель-Ирхуд 1 – Шапель-о-Сен – лицевой отдел (1 случай); Джебель-Ирхуд 1 –
Петралона – лицевой отдел (1 случай); Рабат – Мауэр – нижняя челюсть (1 случай).
8.
Выявлено
тяготение архаичного сапиенса Африки и кроманьонцев Европы. Омо1 – Оберкассель
– мозговой отдел (2 случая).
9.
Выявлено
тяготение кроманьонцев Африки к кроманьонцам Европы. Фиш-Хук – Костенки 14 –
череп без нижней челюсти (1 случай); Фиш-Хук – Оберкассель – мозговой отдел (2
случая); Фиш-Хук – Костенки 14 – лицевой отдел (2 случая); Фиш-Хук – Шанселяд,
Костенки 14, Пшедмости 9 – нижняя челюсть (6 случаев).
10.
Выявлено
тяготение кроманьонцев Африки и неандертальцев Европы. Фиш-Хук – Мустье –
мозговой отдел (1 случай).
Заключение
По нашим результатам,
трансгрессия краниометрических признаков ископаемых гоминид Африки и Азии
выглядит следующим образом:
1.
Африка+Азия; Европа отдельно.
Мозговой отдел
– выявлено при анализе: гейдельбержцы, эректусы. неандертальцы. Лицевой отдел –
эректусы. Кальвариум – австралопитеки.
2.
Африка+Азия+Европа.
Мозговой отдел
- выявлено при анализе: неандертальцы (эректусы, архаичный сапиенс). Кальвариум
– австралопитеки. Лицевой отдел – неандертальцы (гейдельбержцы, эректусы).
Нижняя челюсть – эректусы (неандертальцы). Чаще всего объединение ископаемых
гоминид Африки и Азии встречается при анализе: эректусы → неандертальцы
→ гейдельбержцы (архаичные сапиенсы, австралопитеки). При этом
трансгрессия краниометрических признаков ископаемых гоминид Африки и Европы
выглядит следующим образом: Африка+Европа; Азия отдельно. Мозговой отдел –
выявлено при анализе: неандертальцы (гейдельбержцы). Кальвариум –
австралопитеки. Лицевой отдел – неандертальцы. Нижняя челюсть – эректусы.
3.
Африка+Европа+Азия.
Мозговой отдел
- выявлено при анализе: неандертальцы, гейдельбержцы. Кальвариум –
австралопитеки. Лицевой отдел – эректусы, гейдельбержцы. Нижняя челюсть – эректусы.
неандертальцы. Чаще всего, ископаемых гоминид Африки и Европы объединяют
анализы: неандертальцев, эректусов=гейдельбержцев, австралопитеков.
Таким образом,
последовательность Африка-Азия древнее (?), чем Африка-Европа.
ЛИТЕРАТУРА
4.
Деревянко
А.П. Древнейшие миграции человека в Евразии в раннем палеолите. Новосибирск:
ИАЭТ СО РАН, 2009, С.231.
5.
Дерябин
В.Е., Харитонов В.М. Объективная классификация рода НОМО на основе много мерно
биометрического анализа признаков черепа гоминид//Вопросы антропологии. Вып.91.
М., 2003. С.22-34.
6.
Дробышевский
С.В. «Антропогенез». Учебное пособие для вузов. М., 2009, С.45
7.
Зубов
А.А., Васильев С.В. Первоначальное заселение Европы человеком по данным
антропологии//Доисторический человек: Биологические и социальные аспекты.-
М.:Институт этнологии и антропологии РАН, 2006, С.53-92.
8. Brauer G. Prasapiens-Hypothese oder Afroeuropaische Sapiens
Hypothese//Zeitschrift fur Morhologie und Anthropologie. 1984. B.75. N1. 1-42.
AFRICA -
EUROPE: AN ANALYSIS OF THE DIVERGECE OF
CRANIOMETRIC TRAITS
Kharitonov V. M.
We will continue to analyze the spatial structure of
craniometric recognition of hominid, which covers the three parts of the world
(Africa, Asia, Europe). The results obtained with the help of the canonical
analysis performed on the basis of a computer system HomoBase-Kanoklass,
founded E. V. Deryabin. The material for the work was compiled by craniometry
Pleistocene fossil hominids from Africa, Asia Europe, issued in the form of MS
databases, including measurements of skull 250 hominids. Discusses the results
of the analysis of 82 fossil hominids three parts of the world
(Australopithecus, H.habilis, H. rudolfensis, H.ergaster, H.erecrus,
H.heidelbergensis, H.neanderthalensis, archaic H.sapiens). Canonical analysis
objectively reflects the similarities and differences between the ensembles of
craniometric traits of fossil hominids. Specifically investigated features of
divergence (transgression) craniometric signs of geographical clusters of
Pleistocene hominids in Africa and Europe. In our classification, they are
manifested in variations of geographical groupings of Africa + Europe, Asia -
separately (11.31%) and Africa + Asia + Europe (29, 92%). The magnitude of the
total of the two options - 41.23%. I.e. less than half of all cases studied.
The incidence of cases of transgressions of craniometric characters in hominids
in Africa and Europe is less than that of Africa and Asia, which corresponds to
the expectation of (Europe was populated by migrants, from Africa, Asia later
than in these hominid paleontology). Transgression of craniometric traits of
fossil hominids in Africa and Europe is as follows:
1. Afrika + Europe, Asia separately. The result was obtained in the analysis:
cerebral Department - Neanderthals (H. heidelbergensis), kalvarium -
Australopithecus, the front office – Neanderthals, lower jaw – H.erectus.
2.Afrika + Europe + Asia. The result was obtained in the analysis: cerebral Department – Neanderthals, H.
heidelbergensis , kalvarium - Australopithecus, the front office – H.erectus,
H.Heidelbergensis , the lower jaw – H.erectus, Neanderthals. Most often, the
resources hominid Africa and Europe combined analysis: Neanderthals, H.erectus,
H. heidelbergensis, Australopithecus. Option spatial arrangement of
geographical groupings of fossil hominids Africa + Europe, Asia separately
studied in the canonical set of analytic call rare. He revealed in the study of
hominid lower, middle and upper Pleistocene. One can only assume that the
transgression of the signs of the brain of the skull in hominids in Africa and
Europe is expressed in more cases than the visceral parts of the skull
(actually the front section of the skull, lower jaw). Option Africa + Europe +
Asia - is found in aggregate graphs of fossil hominids, obtained by canonical
analysis, a greater number than version - Africa + Europe, Asia separately. On
the frequency of transgression recognized manifestation of magnitude hominid
skull in Africa and Europe of the skull are as follows: brain department, the
lower jaw →front Department
→ kalvarium. Transgression signs on
display at a frequency of hominids in Africa and Europe is more common in the
following cases: brain section (lower - upper Pleistocene), lower jaw
(lower-upper-price Pleistocene) → front section (lower-upper Pleistocene)
→ kalvarium (Eopleistocene -Lower Pleistocene). According to the
results of phylogenetic relationships of African and Asian hominids-ray appear
to be more ancient than the African and European.
Key words: anthropology (physical anthropology), paleoanthropology,
hominid African, hominids in Europe, signs of divergence
НИИ и Музей антропологии МГУ, Москва
Institute
and Museum of Anthropology, MSU, Moscow
Поступила в редакцию 19.06.2012.