Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 11. -
Вып. 3. - 2012. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/TITL-35.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/cont.htm
УДК 572. 1/.4
ГЕОГРАФИЧЕСКИЕ
АСПЕКТЫ ДИВЕРГЕНЦИИ КРАНИОМЕТРИЧЕСКИХ ПРИЗНАКОВ: АФРИКАНСКИЙ МАТЕРИК – АЗИЯ
Ó 2012 г. Харитонов В. М.
В работе исследовалась дивергенция (трансгрессия) краниометрических
признаков гоминид Африки и Азии при аллопатрического
видообразовании. Проанализированы около 190 графиков канонического анализа
черепных размеров ископаемых гоминид. Материал составляют краниометрические
признаки 250 ископаемых гоминид плейстоцена Африки, Азии и Европы. Варианты
взаимного положения географических групп гоминид: 1) Европа+Азия; Африка
отдельно- 33,94%; 2) Африка+Азия+Европа - 29,92%; 3)Африка+Азия; Европа
отдельно -14,62%; 4) Африка+Европа; Азия отдельно- 11,31%. 4)Африка отдельно;
Азия отдельно; Европа отдельно - 10,22%. Совокупные связи гоминид Африки и Азии
- 44,54%, у гоминид Африки и Европы -
40,69%, а гоминид Азии и Европы - 63,86%. Дивергенция (трансгрессия)
признаков гоминид были обусловлены в разной степени различными отделами черепа. Анализ варианта «Африка+Азия; Европа отдельно» показал, что
не существует выраженной трансгрессии признаков мозгового отдела черепа у гоминид
Африки и Азии, это является свидетельством
эволюционной дивергенции с уровня гоминид нижнего и среднего
плейстоцена. Слабовыраженная дивергенция признаков кальвариума выявлена с
уровня австралопитеков и эректусов. По признакам лицевого отдела дивергенция
гоминид Европы и Азии не выражена, а
трансгрессия, в связи с эволюционной преемственностью, поддержана гоминидами
нижнего и среднего плейстоцена. Анализ
варианта «Африка+Азия+Европа»
выявил трансгрессию признаков мозгового отдела черепа у гоминид трех частей
света, часто наблюдающуюся у гоминид нижнего, среднего и верхнего плейстоцена.
Дивергенция указанных признаков мозгового отдела у гоминид Африки и Азии
выражена слабо. С еще более древнего уровня (с австралопитеков) существует
трансгрессия признаков кальвариума. Она же у признаков висцерального отдела черепа,
обусловленная эволюционной преемственностью, существует у гоминид трех частей
света, начиная с ранних Homo и эректусов. Канонический анализ краниометрических
признаков позволяет выделить филогенетические связи связи африканских
ископаемых гоминид в диапазоне H. rudolphensis – H. heidelbergensis и азиатских гоминид в
диапазоне H.erectus – архаичный H.sapiens.Исходя из моногенизма рода Homo, можно считать, что
дивергенция и трансгрессия краниометрических признаков гоминид Африки и Азии
предопределены различными
продолжительностью и глубиной эволюционного процесса, приведшего к изучаемому
состоянию морфологических признаков. Явление дивергенции связано с продолжительной
и глубокой эволюционной перестройкой признаков черепа, а трансгрессия – с
относительно недавней, когда филогенетическое родство гоминид достаточно
проявлено.
Ключевые слова: антропология (физическая
антропология), палеоантропология, гоминиды Африки, гоминиды Азии, дивергенция
признаков
Происхождение сапиенса во
второй половине XIX века рассматривалось в границах Европейского
континента. До 1921 года находок неандертальского человека за пределами Европы
не было известно. Данное обстоятельство было учтено создателями теории неандертальского
звена [Рогинский,1969]. Теория «широкого моноцентризма» предполагает
становление сапиенса на широкой территории – Южная Азия, Передняя Азия.
Восточное Средиземноморье, Восточная Африка [Рогинский,1969]. В конце 1924 г. с
открытием первого австралопитека в Африке указанный континент был включен в
состав доисторической ойкумены.
По мере накопления находок
ископаемых гоминид, уточнения геологического возраста ископаемых гоминид
стадиальная модель антропогенеза становится не соответствующей действительности. Мы имеем в виду
представления о том, что во всех частях света в каждый отрезок времени
существовал лишь один вид гоминид, а они последовательно образовали линейную
филогению. Наши представления о миграциях гоминид позволяют связать
протоисторию гоминид Африки, Европы и Азии в одно целое, что соответствует
сетчатой модели антропогенеза, предполагающей модусы дивергенции, параллелизмы
в эволюции признаков и конвергенцию. Данный подход ассоциируется с мнением Я.
Я. Рогинского и В. П. Якимова о том, что неравномерность исторического развития
гоминид связана с различием в условиях хозяйствования и степени изолированности
разных групп [Рогинский,1969].
Географический аспект в
исследовании ископаемых гоминид не нов. Например, В. П. Алексеев [1978] в своей
монографии рассматривает географические группировки неандертальцев, анализируя
комплексы краниометрических признаков. В. В. Бунак [ 1980] и Е. Н. Хрисанфова
[1999] анализируют морфологический облик ископаемых гоминид по частям света.
Наконец, А. А. Зубов [2004] в своей всеобъемлющей монографии связал гоминид
различных частей света генетическими связями и миграциями. Крайне интересно
изложение им проблемы в другой работе [Зубов,1993].
А. А. Зубовым предлагается модель дифференциации
человечества на разных этапах на одонтологических материалах. У первых
представителей рода Номо-хабилисов отмечен «западный» морфотип верхних резцов
(нет признаков лопатообразной формы), свойственный современным европеоидам и
негроидам Африки. Он характеризует и эректусов Африки, и некоторые
ближневосточные формы (Амуд, Кафзех), а также известные кроманьонские формы
Европы (Сунгирь). В Азии, начиная с древнейших эректусов, преобладает лопатообразная
форма резцов («восточный» морфотип). Она встречается у европейских неандертальцев
и у некоторых ближневосточных гоминид (Схул, Табун), но в западных регионах не
сохраняется позже палеолита. Интересно, что у современных монголоидов есть
одонтоглифические особенности резцов, сближающие их с «западным» типом. А. А.
Зубов делает вывод, что независимого развития западного и восточного
одонтологических стволов не было, за исключением изолированных отрезков
времени. Современный восточный ствол формировался на базе западного за счет примеси
местных азиатских форм, характеризовавшихся классическим типом верхних резцов,
который мы видим у азиатских эректусов. Где и как происходила метисация – в
Азии или в Европе? В западном регионе, как и в восточном, повсеместно
происходили метисационные процессы, приводившие к появлению «синтетических»
морфологических вариантов зубов. Но носителей западного варианта на западе было
больше, и при метисации неандертальцев и кроманьонцев вклад первых (носителей
«восточного» варианта) был заметно менее половины. На востоке носителей
«восточного» первоначального варианта было больше, и в последующих поколениях
он получает перевес. Генетико-автоматические процессы в периоды изоляции надрасовых
стволов привели к концентрации носителей «западного» типа на западе и типа
«восточного» (обновленного за счет метисации) на востоке.
А. А. Зубов [1995] пишет еще
об одной особенности зубов. Имеется в виду гребень эпикристид, соединяющий
протоконид с метаконидом в середине тригонида. Данный признак отчетливо выражен
на зубах некоторых европейских неандертальцев, типичен для гейдельбержцев,
встречается у африканских архантропов и австралопитековых. Эпикристид маркирует
афро-европейскую ветвь эволюции ранних гоминид. Он редок на востоке и в
современном человечестве (при этом чаще всего встречается у негроидов и южных
европеоидов). Смешение сапиенса с европейскими неандертальцами не привело к
увеличению концентрации данного признака в Европе, так как число
взаимодействующих неандертальских групп было невелико. Далее, эта особенность
дерматоглифики сочетается у европейских неандертальцев с восточным типом
устройства верхних резцов. В этом А.А.Зубов видит сочетание западных и восточных
особенностей у среднеевропейских гоминид, следовательно, настаивает на
«сетевидном» характере эволюции рода Homo.
Невозможно в рамках одной
статьи проанализировать в деталях палеоантропологию двух частей света – Африки
и Азии, делая акцент на генетические связи африканских и азиатских гоминид.
Вспомним, что в науке серьезно рассматривалась азиатская прародина человека,
она и была впервые аргументирована. В
Азии были открыты среднемиоценовые гоминоиды – сивапитеки (8-12 млн. лет
назад). Они до открытия гоминоидов в Африке считались предками гоминид. Но в
Азии нет находок прогрессивных гоминоидов в интервале от 8 до 1,5 млн. л.,
тогда как в Африке обнаружены многочисленные австралопитековые. В настоящее
время азиатские сивапитеки считаются предками лишь современных орангутангов.
Географическое
распространение хабилисов ( и других ранних Homo) ограничивается Африканским
континентом. Из находок вне Африки к Homo habilis некоторые авторы относили тех
же гоминид, кого относили и к австралопитекам – Тель-Убейдиа, Моджокерто. Таким
образом, несомненные "самые ранние Homo" обитали только в Африке,
возможно, несколько севернее, чем австралопитеки. Основные местонахождения
располагаются в Восточной Африке; в Южной Африке отнесение тех или иных находок
к рассматриваемым таксонам более спорно, в частности, из-за худших датировок
[Дробышевский, 2009].
В пределах уровня
архантропов одни и те же находки
определяются как Homo ergaster
или обозначаются как Homo erectus. Чаще
всего под термином Homo ergaster понимаются ранние сравнительно массивные
восточноафриканские ранние
гоминиды, несущие почти полный комплекс
признаков архантропов. Существенно то. что Homo ergaster были первыми гоминидами,
вышедшими за пределы Африки. Кстати, первые мигранты из Африки в Евразию имели
видовой статус, который не имеет однозначного определения [Деревянко, 2009].
Настоящей сенсацией в науке
об ископаемом человеке стало открытие нижней челюсти и двух черепных крышек ископаемого
гоминида в Грузии (Дманиси- район г.Тбилиси).
Анализ фауны, датированной началом позднего виллафранка, позволяет считать дманисского человекаодним
из самых ранних гоминид, переселившихся из Африки в Евразию около 1,8 млн лет назад. Морфотип гоминида сочетает в себе черты ранних Homo Африки
(cкорее – H.ergaster) и древнейших гоминид о.Явы , наряду с некоторыми прогрессивными особенностями
черепа и зубной системы. Дманиский человек – древнейший гоминид западной
Евразии, свидетельствующий о первых миграциях гоминид из Африки по ойкумене
[Харитонов, 2007].
В период 1-1,5 млн. лет
назад архантропы заселили южные районы Европы и Азии, вплоть до Индонезии
(первые находки питекантропов были сделаны именно на острове Ява в конце XIX
века голландским врачом Э. Дюбуа); в Австралию и Америку они не попадали. Эти
архантропы представляют собой наиболее массивных представителей древнейших
людей и относятся к виду Homo erectus ( Pithecanthropus erectus). По мнению А.
П. Деревянко [2009], эректусы были широко распространены в Африке, Азии и,
вероятно, в Европе, они существовали в течении около 1,5 млн лет. Поэтому
закономерен вопрос о степени морфологического отличия эректусов различных
частей света.
Со второй волной мигрантов в
Восточную Азию попали, видимо, гейдельбержцы, они не претерпели эволюционного успеха.
50-40 тыс. лет н. сапиенс пришел в Восточную и Юго-Восточную Азию из
Африки.
Примечателен тот факт, что в
промежутке времени примерно от 500-400 до 130 тыс. лет назад на Земле обитало
крайне разнообразное население, имеющее множество переходных черт – примитивных
и более прогрессивных [Дробышевский, 2009]. Разные части ойкумены были населены
людьми, сильно отличными друг от друга. Часто гоминиды из древнейшей половины
этого интервала рассматриваются как Homo erectus, представители второй половины
– ассоциируются с палеоантропами.
Особый промежуточный облик последних
приводил к большим таксономическим проблемам. С.В. Дробышевский [2009]
считает логичным рассматривать гоминид этого пласта вместе под обобщающим
названием Homo heidelbergensis, хотя их морфологическая неоднородность
очевидна. Изолированные от остального мира популяции на Яве очень долго
оставались почти полностью схожими с Homo erectus и иногда выделяются в вид
Pithecanthropus soloensis.
Отмечается факт явного
несходства в уровне продвинутости синхронных гоминид в разных регионах. Так, в
Юго-Восточной Азии, особенно в Индонезии, в период 130-400 тыс. лет назад ещё
очень часты находки, определяемые как Homo erectus; в Китае есть множество
находок, определяемых их исследователями как "архаический Homo
sapiens", хотя их признаки далеко не так сапиентны, как у синхронных им
африканских гоминид. В Европе местами тоже жили довольно архаичные люди, но
преобладали, видимо, Homo heidelbergensis (эти европейские формы иногда
определяются как "атипичные неандертальцы" или
"пресапиенсы"). В Африке в то же время появляются значительно более
сапиентные люди, иногда классифицируемые как Homo helmei, причём их датировки
удревняются даже до 490 тыс. лет назад. Впрочем, большинство датировок для Homo
helmei занимают промежуток от 100 до 200 тыс. лет назад. Примерно со 130-160
тыс. лет назад или даже ранее в Африке и на Ближнем Востоке сапиентные признаки
населения становятся настолько выраженными, что часто говорится о появлении в
это время вида Homo sapiens, однако более внимательный анализ обычно показывает
наличие в их строении не меньшего количества и архаичных признаков
[Дробышевский. 2009].
Некоторые синхронные
европейским неандертальцам популяции Африки и Ближ-него Востока были
значительно больше похожи на современного человека. Многие иссле-дователи даже
относят их к современному виду. У людей из Херто ( Эфиопия), Клазиес Ривер (
Южная Африка), из пещер Схул и Джебел Кафзех (Израиль) и некоторых других
местонахождений имелся подбородочный выступ, затылок был округлый, а череп высокий.
Считается, что размер и форма мозга этих людей почти неотличимы от современного
варианта. Датировки же превышают 100 тысяч лет. Промежуточность облика
некоторых ближневосточных находок (Схул, Табун) интерпретируется трояко: некоторые
считают их за эволюционно промежуточную стадию между неандертальцами и
современными людьми, другие – за особую специализированную группу, третьи – за
метисов; последние две точки зрения имеют несколько лучшую аргументацию, чем
первая [Дробышевский, 2009].
Ближний Восток и Центральная
Азия были заселены людьми двух типов. Одни были больше похожи на неандертальцев
Европы, другие – на прогрессивных палеоантропов Африки и людей из пещер Схул и
Джебел Кафзех. Характерно, что культура всех этих людей была весьма схожей.
На Дальнем Востоке
население, синхронное гейдельбержцам Европы и Африки – до 130 тысяч лет назад,
почти не отличалось от них внешне. В дальнейшем судьба этого населения неясна.
Антропологические находки с Дальнего Востока в промежутке от 130 до 40 тысяч
лет назад неизвестны. Потом там сразу появляются люди полностью со-временного
облика. Объяснение этому феномену многие учёные видят в "гипотезе
вулканической зимы". Вероятно, практически всё население Южной Азии
вымерло в результате этого катаклизма. Хотя есть археологические свидетельства,
что некоторые группы всё же пережили его, например, в 2004 г. был описан особый
вид людей с индонезийского острова Флорес – Homo floresiensis, – живший 95-12
тыс. лет назад и обладавший крайне малым мозгом – около 40 см3, ростом около 1
м, сильно удлинёнными руками, но изготавливавший орудия труда и охотившийся на
местных карликовых слонов-стегодонов. В Европе неандертальцы именно тогда
приобрели свой "классический ледниковый" облик и в борьбе с тяжелейшими
условиями жизни отстали в своём развитии, а Африка, будучи менее прочих
регионов задета катастрофой, стала родиной нового, наиболее прогрессивного вида
– Homo sapiens [Дробышевский. 2009].
Останки первых людей, почти
совсем неотличимых он нас, обнаружены в нескольких африканских местонахождениях
с датировками от 200 до 100 тысяч лет назад – Херто, Клазиес Ривер, Бордер
Кэйв, Синга, Дар-эс-Салтан. Они не имели сильно выступавшего затылка, большого
надбровного валика и обладали при этом очень большим мозгом и выступающим
подбородком. Похожие находки были сделаны на Ближнем Востоке – в пещерах Схул и
Кафзех. С момента около 40-45 тысяч лет назад люди совершенно современного
облика, только несколько более массивные, чем мы, – неоантропы – известны
практи-чески со всей территории ойкумены – из Африки, Европы, Азии и Австралии.
Только Америка была заселена позже – достоверно только 11-12 тысяч лет назад,
хотя некоторые археологи отодвигают это событие даже до 30-40 тысяч лет назад.
В одной из своих работ
[Дерябин, Харитонов, 2003] мы касались пространственных и филогенетических
взаимоотношений плейстоценовых гоминид Африки и Азии, обсуждая одну из
современных моделей появления ископаемого сапиенса и неандертальцев. Результаты
канонического анализа краниометрических признаков ископаемых гоминид, по-нашему
мнению, позволяют апробировать "афро-европейскую гипотезу" [Brauer
G., 1984]. В частности, рассматривался этап "афро-азиатский
(ближневосточный). В результате проведенного
статистического анализа выяснилось, что
гейдельбержец Брокен-Хилл (0,3 млн.лет)
и питекантроп Олдувей (0,36 млн.лет) не ближе к группе архаичных сапиенсов Схул
чем к синантропам по признакам мозгового отдела черепа. Отличия гейдельбержцев
Брокен-Хилл и Салданьи от раннего Зуттие и группы архаичных сапиенсов Схул -
соотносимы с различиями схульцев и азиатских H.erectus, "финальными"
erectus и азиатским неандертальцем Мапа. Африканский Брокен-Хилл далее от
ближневосточного сапиенса Табун, чем от поздних западноевропейских
неандертальцев. При этом африканский архаичный сапиенс Омо II близок к
ближневосточному сапиенсу Схул IY, а
Омо I к ближневосточному
сапиенсу Кафзех. Лишь по единичным признакам питекантроп Олдувай, гейдельбержцы Ньярасса и Брокен-Хилл ближе к
членам группы ископаемых сапиенсов Схул чем к азиатским H.erectus.
По признакам лицевого отдела
черепа расположение африканского гейдельбержца Брокен-Хилл, европейского гейдельбержца
Петралоны и ближневосточных архаичных sapiens выглядит сложными: Брокен-Хилл
ближе к европейской Петралоне, чем к Схул IY и Табун. Далее. Различие
Брокен-Хилла и кроманьонцев аналогично
тому, что констатировано между схульцами и европейскими неандертальцами.
Гейдельбержец Брокен-Хилл отличен от архаичных сапиенсов Ближнего Востока Схул
Y,IY,IX, неандертальца Амуд более чем
от европейских неандертальцев. О признаках нижней челюсти можно сказать
немного: европейский гейдельбержец Араго XIII ближе к атлантропу, чем к членам
группы архаичных сапиенсов Схул-Табун. В указанной работе был сделан вывод:
конкретные связи между ископаемыми гоминидами т.н. афро-ближневосточного вектора достаточно сложны.
В своей работе мы
руководствовались тем, что дивергенция форм, по мнению И. И. Шмальгаузена
[1969], лежит в основе эволюционного процесса. Возможность дивергенции обусловлена
спецификой реакций на изменение среды, при этом зависит от наследственных
особенностей организмов. Длительность процесса дивергенции зависит от степени
специализации форм и имеет свои пределы. В общем, эволюция организмов идет под
знаком дифференциации и расхождения признаков.
Материал и
методы исследования
В нашей работе сделана
попытка выявить пространственную структуру краниометрических признаков гоминид,
охватывающую три части света (Африку, Азию, Европу) с помощью результатов канонического
анализа, проведенного на основе системы HomoBase-Kanoklass, созданной профессором
Е.В.Дерябина. Канонический анализ объективно отражает сходство и различие
ансамблей краниометрических признаков ископаемых гоминид.
Материалом для работы послужили
данные по краниометрии ископаемых гоминид плейстоцена Африки, Азии Европы,
оформленные в виде РС базы данных, разработанной д.б.н. В.Е.Дерябиным.
Указанная база включает измерения черепа 250 гоминид. В настоящей работе
обсуждены результаты канонического анализа 82 ископаемых гоминид трех частей
света. Имеются в виду: а) австралопитеки – 3 вида; б) H. habilis-1, в) H. rudolfensis-1,
г) H. ergaster- 1, д) H. erecrus-29, е) H. heidelbergensis-12, ж) H. neanderthalensis-23, з) архаичный H. sapiens-
12.
При проведении настоящей
работы использовалась модификация канонического анализа [Key, Jantz, 1990,
1990а], когда рассматривалась не межвыборочная, а общая вариация, наблюдаемая в
совокупности индивидуальных объектов, разного таксономического статуса. В ней,
очевидно, присутствуют как межиндивидуальный внутритаксонный (внутригрупповой)
компонент изменчивости, так и - собственно межгрупповая межтаксонная часть
вариации, на выявление и описание которой направлен анализ.
Средствами СУБД FoxPro 2.5
(версия для DOS) была построена база данных HOMOBASE, в качестве содержательных
полей включавшая значения 49 классических размеров нейрокраниума, лицевой зоны
кальвариума и нижней челюсти. Ключевые поля для формирования запросов из базы
данных включали информацию: по континенту и его региону, где была сделана
находка, полу, возрасту, эпохе и предположительному видовому отнесению. Для
проведения анализа данных база данных HOMOBASE была сопряжена со специально
разработанной для этих целей компьютерной программой КАНОКЛАС 6.5.
Следует заметить, что
никакой предварительной разбивки гоминид для таксономического анализа по
признаку эволюционного уровня не проводилось. Прогнозируемый эволюционный
статус и древность гоминида обсуждались в связи с результатами проведенного
статистического анализа. Обозначенные условия работы, по-нашему мнению, обусловили
реальность ее выводов. Относительность результатов применения кластерного
анализа для выработки объективной классификации Hominidae в нашей работе
обусловлена выборочным анализом ископаемых объектов, фрагментарностью
материала, неравной представленности в анализе признаков традиционных отделов
черепа [Дерябин, Харитонов, 2003].
Результаты и
обсуждение
Предметом нашего изучения
явилась дивергенция/трансгрессия краниометрических признаков гоминид,
возникающая при видообразовании, аллопатрическом, пространственном. Степень
дивергенции/трансгрессии краниометрических признаков, выявляемой с помощью
канонического анализа, зависит от многих объективных причин, как-то несходства
сред обитания, наследственных особенностей, специфики происходящего эволюционного
процесса и т.д. Основной задачей исследования является соотнесение картины дивергенции
краниометрических признаков гоминид с реконструированной картиной их географической
дивергенции. Естественно, необходимо учитывать, что изучаемая дивергенция
признаков ограничивается процессами конвергенции признаков и параллельного
развития, которые несомненно имели место в антропогенезе.
Взаимное положение
географических групп ископаемых гоминид, как нам кажется, на графиках отражает
историю их эволюционных взаимоотношений. Двигаясь по оси времени от древности к современности, мы видим
пространственно-временную динамику взаимоотношений гоминид в трех частях света:
Африке, Азии и Европе. Это важно, так как чаще всего в анализе эволюции гоминид
во внимание принимается лишь ось времени. При этом, мы согласны с С. В.
Дробышевским [2000] в том, что географический критерий является второстепенным
в сравнении с морфологическим, когда речь идет о разграничении фенонов
ископаемых гоминид в связи с установленными фактами миграций ископаемых гоминид
и возможностью их сосуществования на определенных территориях.
1. Общий обзор географического анализа графиков канонического анализа
ископаемых гоминид.
В работе проанализированы
191 график канонического анализа краниометрических признаков ископаемых гоминид
и их предшественников( австралопитековых). Первоначальный анализ показал, что
ситуации пространственного взаимного положения группировок ископаемых гоминид
на материале краниометрических признаков были распределены по возрастающей между
следующими вариантами:
а) Трансгрессия гоминид
Европы и Азии; Африка отдельна; 33, 94% (93 случая),
б)
Трансгрессия гоминид Африки, Азии и Европы; 29,92% (82 случая),
в) Трансгрессия
гоминид Африки и Азии; Европа отдельно; 14,62% (40 случаев),
г)
Трансгрессия гоминид Африки и Европы; Азия отдельно. 11,31% (31 случай),
д)
Отсутствие трансгрессии гоминид Африки, Европы и Азии; 10,22%(28 случаев).
Итого-274
случая. Итоговая величина числа случаев, которая превышает количество графиков
канонического анализа, объясняется тем, что в случаи отсутствия гоминид одной
части света, мы присоединяли данный вариант ко всем возможным.
Простая
статистика встречаемости вариантов
взаимного положения частей света по признакам трансрессии и дивергенции наводит
на следующие мысли. Вариант «д» - очень редок, так как он предполагает
независимые взаимоотношения гоминид в границах каждой из трех частей света.
Анализ всей изученной
совокупности случаев трансгрессии географических групп гоминид позволяет
заключить следующее. Совокупные связи гоминид Африки и Азии могут быть оценены
в 44,54%, они же у гоминид Африки и Европы в 40,69%, т.е. несколько меньше, а
объем связей гоминид Азии и Европы в 63,86%.
Явления трансгрессии и дивергенции признаков черепа гоминид были
выявлены с помощью канонического
анализа диаметров следующих отделов черепа.
1. Европа+Азия; Африка
отдельно (МО=43; МО+ЛО=3; ЛО=21; НЧ=26)
2. Африка+Азия+Европа
(МО=38; МО+ЛО=5; ЛО=14; НЧ=25)
3. Африка+Азия; Европа
отдельно (МО=32; МО+ЛО=3; ЛО=3; НЧ=1)
4. Африка; Азия; Европа –
все отдельно (МО=20; МО+ЛО=1; ЛО=3; НЧ=4)
5. Африка+Европа; Азия
отдельно (МО=29; ЛО=1; НЧ=1)(где МО-мозговой отдел; ЛО-лицевой отдел; МО+ЛО-
кальвариум; НЧ- нижняя челюсть).
Мы видим, что в первом и
втором случаи (наиболее часто встречаемом) повышена частота анализов нижней
челюсти, можно говорить также о повышенной частоте встречаемости анализов
признаков мозгового и лицевого отделов черепа.
2. Дивергенция краниометрических признаков у ископаемых гоминид Африки,
Азии и Европы.
Особняком от других стоит
вариант взаимоотношения географических группировок ископаемых гоминид – Африка
отдельно; Азия отдельно; Европа отдельно (вариант «в»). Он встречается в 10,22
% случаев. Видимо, он отражает независимые эволюционные взаимоотношения, в том
числе дивергенцию признаков черепа ископаемых гоминид в указанных частях света.
Данные изменения обратные трансгрессии краниометрических признаков,
обусловленной эволюционной преемственностью.
Распределение случаев
дивергенции географических групп гоминид по отделам черепа выглядит так.
Мозговой отдел – 2 случая
(рис.1а) (Европа отдельно; Азия отдельно – 18 случаев).
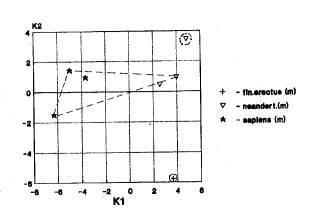
На рис.1а мы видим
разнесенное в пространстве положение африканских гейдельбержцев, азиатских
неандертальцев, а также европейских неандертальцев и кроманьонцев.
Кальвариум (мозговой отдел +
лицевой отдел) – 1 случай (рис.1б).
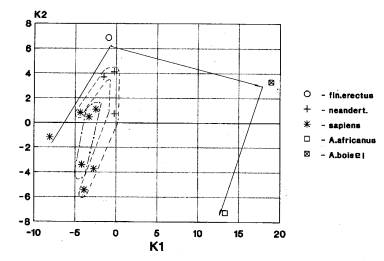
На рис.1б мы видим не перекрывающееся положение африканских австралопитеков и гейдельбержцев, европейских неандертальцев и кроманьонцев, а также азиатских кроманьонцев.
Лицевой отдел – 3 случая
(рис.1в).
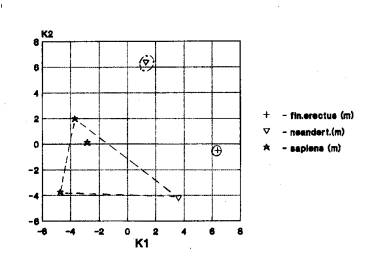
На рис.1в мы видим взаимное
положение африканского гейдельбержца, азиатского кроманьонца и европейских
неандертальцев и кроманьонцев.
Нижняя челюсть – 4 случая (рис.1г).
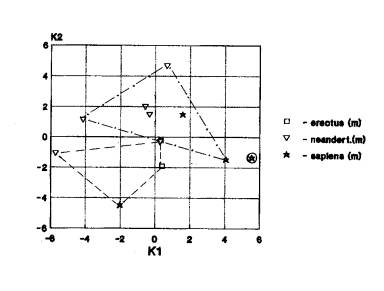
На рис.1г мы видим
пространственную разобщенность кроманьонцев Африки, эректусов, неандертальцев,
архаичных сапиенсов и кроманьонцев Азии и эректусов. неандертальцев и
кроманьонцев Европы.
Анализ дивергенции признаков
черепа у ископаемых гоминид трех частей света по отделам черепа показал, что
расхождение сильнее выражено у признаков нижней челюсти, меньше – у признаков
лицевого отдела и менее всего у признаков мозгового отдела и кальвариума. Возможная трансгрессия признаков
черепа у гоминид трех частей света имеет обратный градиент: Кальвариум →
Мозговой отдел→ Лицевой отдел → Нижняя челюсть.
Указанное эволюционное
явление поддерживают у указанных географических группировок следующие виды.
Дивергенция по признакам мозгового черепа наблюдается у гейдельбержцев,
висцерального черепа – австралопитеков, лицевого отдела – чаще у неандертальцев,
нижней челюсти – питекантропов и неандертальцев. Т.е. явление дивергенции
проявляется чаще у ископаемых гоминид нижнего и среднего плейстоцена.
3. Обсуждая вопрос о взаимоотношениях гоминид Африки и Азии,
мы видим, что величина их выражена в сумме встречаемости вариантов «б» и «в»,
т.е. 29,92% +14,65 =44,54% (меньше половины изученных случаев).
Проанализируем каждый
вариант (в и б) отдельно.
Вариант «в». Африка+Азия;
Европа – отдельно.
Указанный вариант
пространственного расположения географических группировок гоминид получен при
каноническом анализе следующих отделов черепа: мозговой отдел - 32 случая; кальвариум - 3 случая; лицевой отдел – 3 случая; нижняя
челюсть – 1 случай.
Первоначальный анализ
вклада разных отделов черепа гоминид в
явление трансгрессии признаков позволил предположить, что последняя выражена у
мозгового отдела (т.е. дивергенция проявляется минимально). Дивергенция
признаков более выражена по признакам лицевого черепа (включая нижнюю челюсть и
всего черепа без нижней челюсти). Т.е. наибольшей эволюционной перестройке у
гоминид Африки и Азии подвергся висцеральный череп. Относительный вклад отделов
черепа в явление трансгрессии признаков показал, что филогенетические связи
гоминид указанных частей света прежде всего проявляются в мозговом отделе. Надо
заметить, что совсем изолированное положение европейских гоминид по отношению к
африканским и азиатским наблюдается крайне редко.
Вариант «б».
Африка+Азия+Европа.
Указанный вариант
пространственного расположения географических групп гоминид получен при по
результатам канонического анализа следующих отделов черепа: мозговой отдел-38
случаев; нижняя челюсть-25 случаев; лицевой отдел-14 случаев; кальвариум - 5
случаев.
Трансгрессия признаков, если
ориентироваться на первоначальный общий анализ встречаемости вариантов
взаимного положения географических групп гоминид, более всего выражена у
мозгового отдела гоминид трех частей света, далее следуют признаки нижней
челюсти, далее – лицевой отдел; далее –
кальвариум. Дивергенция признаков будет выражена обратным образом.
Наибольшей перестройке в эволюции, что отражается в дивергенции признаков
подвергся висцеральный череп.
Проведем подробный анализ
картины трансрессии и дивергенции краниометрических признаков у гоминид Африки
и Азии с учетом эволюционного уровня гоминид, вошедших в него и, выделяя
количество бесспорных и возможных
результатов анализов.
Вариант «в». Африка+Азия;
Европа – отдельно.
Мозговой отдел. Выявлены следующие варианты взаимного расположения географических группировок: 1. Африка+Азия; Европа отдельно (2 случая) (рис.1д), 2. Африка+Азия…. Азия+Европа (4 случаев), 3. Азия…. Европа (26 случаев). В первом случаи уровень определен эректусами и гейдельбержцами. Во втором случаи вариант поддержан в равной степени гейдельбержцами. В третьем случаи вариант поддержан эректусами → неандертальцами → гейдельбержцами →австралопитеками.
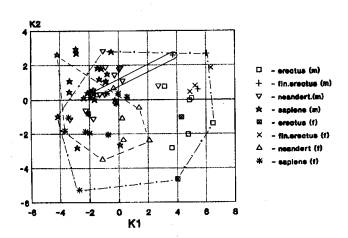
На рис.1д мы наблюдаем сдвиг
области значений европейских неандертальцев и кроманьоцев по отношению к африканским
гейдельбержцам и кроманьонцам, а также к азиатским эректусам,гейдельбержцам,
неандертальцам,архаичным сапиенсам и кроманьонцам, которые показывают полное
совмещение в пространстве своих значений.
Лицевой отдел. Выявлены следующие варианты взаимного положения географических группировок: Африка+Азия… Азия+Европа (3 случая) (рис.1е). Преобладает трансгрессия признаков лицевого отдела у эректуса . Европа не остается полностью в стороне, она контактирует с Азией.
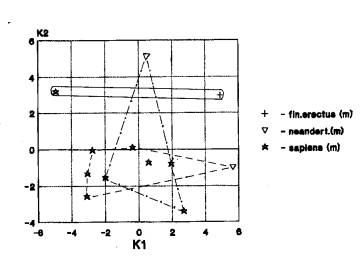
На рис. 1е мы видим попарное
наложение областей значений африканских гоминид(гейдельбержцы и кроманьонцы) и
азиатских (архаичные сапиенсы и кроманьонцы), с одной стороны, азиатских и европейских (неандертальцы и
кроманьонцы), с другой.
Кальвариум. Выявлены следующие варианты взаимного расположения географических группировой анализируемых гоминид. 1. Африка+Азия… Азия+Европа (1 случай), 2. Африка+Азия; Европа отдельно (2 случая)(рис. 1ж). В том случаи, когда в анализ входят австралопитековые, то наблюдается второй вариант.
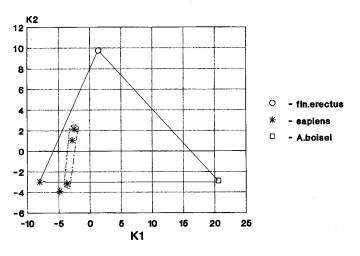
На рис.1ж мы видим
трансгрессию значений африканских гоминид (австралопитеки, гейдельбержцы,
кроманьонцы) и азиатских (кроманьонцы).
Нижняя челюсть. Наблюдается
один вариант – Азия…Европа (1 случай). Он выявлен при анализе эректусов.
Подводя итоги анализа
варианта пространственного расположения географических группировок Африка+Азия;
Европа отдельно, мы видим, что многочисленные случаи с признаками мозгового
отдела не показывают убедительной трансгрессии признаков у гоминид Африки и
Азии (2+4 случаев). Это явление можно рассматривать как свидетельство
существования и эволюционной дивергенции. Зато изолированность по ним гоминид
Азии и Европы убедительна (26 случаев). Она проявляется в анализах с уровня
гоминид нижнего и среднего плейстоцена, т.е. дивергенция здесь налицо.
По признакам кальвариума (череп без нижней челюсти) трансгрессия
признаков поддерживается австралопитеками и эректусами. Здесь дивергенция
выражена слабо.
По признакам лицевого отдела
дивергенция гоминид Европы и Азии не выражена (?), а трансгрессия (эволюционная
преемственность) поддержана гоминидами нижнего и верхнего плейстоцена.
О дивергенции признаков
нижней челюсти судить трудно. Может быть, она у гоминид нижнего плейстоцена
отсуствует?
Вариант «б». Африка+Азия +
Европа.
Мозговой отдел. Выявлены следующие варианты взаимного расположения географических группировой гоминид: 1. Африка+Азия+Европа (21 случай) (рис.1з), 2. Азия+Европа… (17 случаев). В первом случаи уровень анализа начинается с неандертальцев, менее – с архантропов, менее – гейдельбержцев. Во втором случаи указанная конфигурация группировок поддерживается архаичным сапиенсом, далее идут неандертальцы, архантропы и гейдельбержцы.
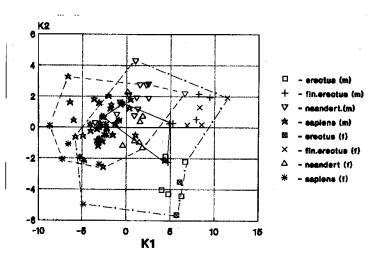
Рис.1з. Мы видим
трансгрессию значений гоминид Африки (гейдельбержцев, кроманьонцев), Азии(
питекантропы, гейдельбержцы, архаичные сапиенсы, кроманьонцы), Европы
(неандертальцы, кроманьонцы).
Кальвариум. Выявлен только
один вариант взаимного расположения географических группировок:
Африка+Азия+Европа (5 случаев). Чаще всего он получается при анализе австралопитековых
разных видов (может быть, в связи с большим размахом присущей им межгрупповой
изменчивости).
Лицевой отдел. Выявлены следующие варианты взаимного положения географических группировок: 1. Африка+Азия+Европа (10 случаев)(рис.1и). 2. Африка+Азия…Азия+Европа (4 случая). Первый вариант получен при анализе с уровня неандертальцев, в меньшей степени – гейдельбержцев, еще меньше – эректусов.
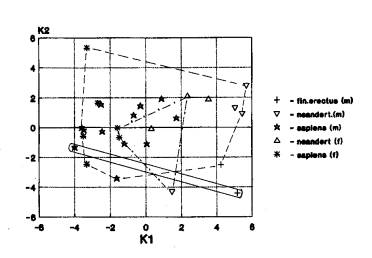
Рис.1и. Мы видим
трансгрессию признаков лицевого отдела у гоминид Африки (гейдельбержцы,
кроманьоцы), Азии (архаиный сапиенс, кроманьоцы) и Европы(гейдельбержцы,
неандертальцы, кроманьонцы)
Нижняя челюсть. Наблюдается и преобладает один вариант – Африка+Азия+Европа (24 случая)(рис.1к). Он выявляется прежде всего при анализе эректусов, в меньшей степени- неандертальцев, гейдельбержцев, хабилисов.
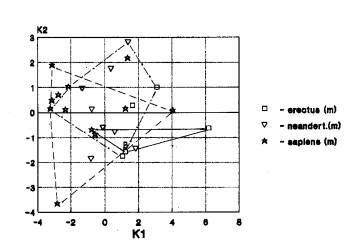
На рис.1к мы видим
трансгрессию признаков гоминид Африки(хабилис, питекантропы, кроманьонцы), Азии
(питекантропы,неандертальцы, архаичный сапиенс, кроманьонцы), Европы
(неандертальцы, кроманьонцы).
Подводя итоги анализа варианта пространственного расположения
географических группировок Африка+Азия+Европа, можно заключить, что
трансгрессию признаков мозгового отдела черепа у гоминид трех частей света мы
наблюдаем в 50% случаев, она выявлена при анализе гоминид нижнего, среднего,
верхнего плейстоцена, начиная с H.erectus. Можно предположить, что
еще чаще встречается за счет варианта трансгрессии, в котором объединены
признаки мозгового отдела гоминид Азии и Европы, начиная с того же уровня.
Видимо, можно говорить, что дивергенция признаков мозгового отдела у гоминид Африки
и Азии выражена слабо. С еще более низкого уровня (с австралопитеков) наблюдается
трансгрессия признаков у черепа без нижней челюсти (мозговой и лицевой отделы
вместе). Трансгрессия признаков, обусловленная эволюционной преемственностью,
видна у лицевого отдела черепа гоминид трех частей света, начиная с ранних Homo и
нижнеплейстоценовых гоминид. Трансгрессия признаков нижней челюсти преобладает
у гоминид Африки и Азии, начиная с H.erectus.
4. Межрегиональные связи
стадиальных групп ископаемых гоминид Африки и Азии.
Эволюция гоминид
прослеживается между конкретными
стадиальными и внутристадиальными группами ископаемых гоминид. Соотношение
верхнеплейстоценовых гоминид Африки и Азии особенно интересно, так как
маркирует возможные филогенетические связи ископаемых гоминид Африки и Азии, а
также показывает на генезис сапиенса, совпадающий в географическом аспекте с
миграциями гоминид из Африканского материка в Азию, а затем в Европу.
Анализ 191 графика канонического анализа краниометрических
признаков ископаемых гоминид Африки и Азии продемонстрировал следующие варианты
морфологической близости.
1.
Выявлено
тяготение гейдельбержцев Африки и архаичных сапиенсов Ближнего Востока, а
именно: а)Бодо, Броккен-Хилл – Схул IY, Схул V –
признаки мозгового и лицевого отделов черепа (5 случаев); б)Бодо, Броккен-Хилл
– Схул IY, Схул Y, Схул IX – мозговой отдел (5
случаев); в)Брокен-Хилл – Схул IY, Схул IX–
лицевой отдел (5 случаев); г)Омо II – Схул IX –
мозговой отдел (1 случай).
2.
Выявлена
близость африканских гейдельбержцев и
азиатским архантропам разной видовой принадлежности: а)Рабат – синантроп –
нижняя челюсть (5 случаев); б)Броккен-Хилл – синантроп, питекантроп – мозговой
отдел (5 случаев); в)Ньярасси – синантроп, питекантроп – мозговой отдел (4
случая).
3.
Выявлено
тяготение африканских архантропов к азиатским палеоантропам, а именно: а)
Атлантроп III – Шанидар II – нижняя челюсть (1
случай); б) Атлантроп II – Амуд I –
нижняя челюсть (1 случай).
4.
Выявлено
тяготение H.rudolphensis Африки к азиатскому
архантропу: а) H. rudolphensis – питекантроп – мозговой
отдел (1 случай).
Таким образом,
прослеживаются связи африканских ископаемых гоминид в диапазоне H. rudolphensis – H. heidelbergensis и азиатских гоминид в
диапазоне H. erectus – архаичный H. sapiens.
Заключение
Исходя из моногенизма рода Homo,
можно считать, что дивергенция и трансгрессия краниометрических признаков гоминид
Африки и Азии предопределены различными
продолжительностью и глубиной эволюционного процесса, приведшего к изучаемому
состоянию морфологических признаков. Видимо, дивергенция связана с
продолжительной и глубокой эволюционной перестройкой признаков черепа, а
трансгрессия – с относительно недавней, когда филогенетическое родство гоминид
достаточно проявлено. Проведенный в работе анализ показал на продолжительный и
глубокий характер эволюционного процесса ископаемых гоминид Африки и Азии.
ЛИТЕРАТУРА
1.
Алексеев
В.П. Палеоантропология земного шара и формирование человеческих рас. Палеолит.
М.: Наука, 1978, С.283.
2.
Бунак
В.В. Род Homo, его возникновение и
последующая эволюция. М.: Наука, 1980, С.328.
3.
Деревянко
А.П. Древнейшие миграции человека в Евразии в раннем палеолите. Новосибирск,
2009, С.231.
4.
Дерябин В.Е., Харитонов В.М. Объективная
классификация рода Homo
на основе многомерного биометрического анализа признаков черепа
гоминид//Вопросы антропологии, вып.91. М.,2003. С.22-34.
5.
Дробышевский
С.В. Эволюция краниометрических признаков гоминид (канонический анализ).
М.-Чита, ЗИП Сиб УПК, 2000, С.141.
6.
Дробышевский
С.В. «Антропогенез». Учебное пособие для вузов. М., 2009, С.45
7.
Зубов
А.А. Проблемы внутриродовой систематики рода НОМО в связи с современными
представлениями о биологической дифференциации человечества//Современная
антропология и генетика и проблема рас у человека. М.: Институт этнологии и
антропологии РАН, 1995. С.18-43.
8.
Зубов
А.А. Палеоантропологическая родословная человека. М.: Институт этнологии и
антропологии РАН, 2004, С.552.
9.
Рогинский
Я.Я. Проблемы антропогенеза. М.: Изд-во МГУ,1969, С.262.
10.
Харитонов
В.М. Ископаемые гоминиды Кавказского региона (статистическое исследование)//
Человек в культурной и природной среде (Труды 3 антропологических чтений к
75-летию со дня рождения академика В.П.Алексеева. Москва, 15-17 ноября 2004
г.). М.,Наука,2007, С.108-115.
11.
Хрисанфова
Е.Н., Перевозчиков И.В. Антропология. М.: Изд-во МГУ,1999. С. 397.
12.
Шмальгаузен
И.И. Проблемы дарвинизма. Л: Наука, 1969. С.493.
13.
Brauer G. Prasapiens-Hypothese oder
Afroeuropaische Sapiens Hypothese//Zeitschrift fur Morhologie und
Anthropologie. 1984. B.75. N 1. P. 1-42.
Рисунки
1а- 1к – Взаимное расположение областей значений
краниометрических значений гоминид Африки, Азии и Европы
Обозначение:
ــــــ область значений
гоминид Африки,
- - - - область значений гоминид Европы,
- · - · - область значений гоминид Азии
GEOGRAPHICAL ASPECTS DIVERGENCE CRANIOMETRIC SIGNS:
AFRICAN CONTINENT – ASIA
Kharitonov
V. M.
We investigated the divergence (transgression) craniometric traits of
minid, Africa and Asia during allopatric speciation. Analyzed about 190 plots
of the canonical analysis of cranial dimensions of fossil hominids. Material
form craniometric features 250 of fossil hominids Pleistocene of Africa, Asia
and Europe. Variants of the relative position of the hominid geographic groups:
1) Europe + Asia, Africa alone, 33.94%, and 2) Africa + Asia + Europe - 29.92%
3) Africa + Asia, Europe alone -14.62%, and 4) Africa + Europe, Asia, 11.31%
separately. 4) separately Africa, Asia alone, Europe separately - 10.22%. The
total due hominids in Africa and Asia - 44.54%, the hominids in Africa and
Europe - 40.69%, and hominids in Asia and Europe - 63.86%. Divergence (transgression),
signs of hominids were due to different degrees by different things, from the
skull. Analysis options "Africa + Asia, Europe alone" has shown that
there is no transgression's expression of symptoms of cerebral skull of
hominids in Africa and Asia, it is a testament to the evolutionary divergence
of hominids from the level of the lower and middle-Pleistocene. Mild signs of
divergence kalvarium detected at the level of Australopithecus and erectus. On
the grounds of the facial part of the divergence of hominids in Europe and Asia
is not expressed, and transgression, in connection with the evolutionary
continuity, supported by the hominids of the lower and middle Pleistocene.
Analysis options "Africa + Asia + Europe" showed signs of
transgression of the brain-wave of the hominid skull in three parts of the
world, often seen in hominids a lower, middle and upper Pleistocene. The
divergence of these signs of brain-case in hominids in Africa and Asia is weak.
With an even more ancient level (with the Australopithecus) there is a
transgression of the signs kalvariuma. It also features a visceral-General of
the skull caused by the evolutionary continuity, there is a hominid three parts
of the world, starting with the earliest Homo erectus, and Canonical analysis
of craniometric features allows you to emphasize phylogeny-optical
communication connection African fossil hominids in the range of H.
rudolphensis - H. heidelbergensis and Asian hominids in the range H.erectus -
archaic H.sapiens. Based monogenism the genus Homo, we can assume that the divergence
and the transgression of the craniometric evidence of hominids in Africa and
Asia by various predetermined duration and depth of the evolutionary process
that led to the studied of states of morphological characters. The phenomenon
of divergence due to the long and deep evolutionary rearrangement of features
of the skull, and the transgression - a relatively recent, when the
phylogenetic relationship of hominids rather exposed.