Математическая морфология.
Электронный математический и
медико-биологический журнал. - Т. 11. -
Вып. 3. - 2012. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/TITL-35.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-35-html/cont.htm
УДК 572. 1/.4
ДИВЕРГЕНЦИЯ
ПРИЗНАКОВ ЧЕРЕПА ИСКОПАЕМЫХ ГОМИНИД АЗИИ И ЕВРОПЫ
Ó
2012 г. Харитонов В. М.
Исследовалась дивергенция
(трансгрессия) краниометрических признаков гоминид при аллопатрического
видообразовании. Проанализированы около
190 графиков канонического анализа черепных размеров ископаемых гоминид.
Материал составляют краниометрические признаки 250 ископаемых гоминид
плейстоцена Африки, Азии и Европы. Варианты
взаимного положения географических групп гоминид: 1) Европа+Азия; Африка
отдельно - 33,94%; 2) Африка+Азия+Европа - 29,92%; 3) Африка+Азия; Европа отдельно
-14,62%; 4) Африка+Европа; Азия отдельно- 11,31%. 4) Африка отдельно; Азия
отдель-но; Европа отдельно - 10,22%. Совокупные связи гоминид Африки и Азии -
44,54%, у гоминид Африки и Европы -
40,69%, а гоминид Азии и Европы - 63,86%. Дивергенция (трансгрессия)
признаков гоминид были обусловлены в разной степени различными отделами черепа.
Выделены следующие варианты взаимного положения географических кластеров гоминид.
Вариант – Азия+Европа; Африка отдельно. Ископаемые гоминиды Азии и Европы
демонстрируют трансгрессию признаков в следующих диапазонах времени: а)
мозговой отдел – нижний-верхний плейстоцен или эоплейстоцен-верхний плейстоцен;
б) кальвариум - эоплейстоцен-средний плейстоцен. в) лицевой отдел -
средний-верхний плейстоцен, г) нижняя челюсть - нижний-верхний плейстоцен.
Филогенетические связи ис-копаемых гоминид Азии и Европы охватывают широкий
диапазон – эоплейстоцен - верхний плейстоцен, но чаще нижний - верхний
плейстоцен. Трансгрессия краниометрических признаков ископаемых гоминид Азии и
Европы осуществляется за счет мозгового отдела, реже - нижней
челюсти, лицевого отдела и кальвариума. Чаще всего за счет первых трех
отделов черепа. Вариант - Африка+Азия+Европа. Трансгрессия краниометрических
признаков осуществ-лялась : а) мозговой отдел – нижний-верхний плейстоцен; б)
кальвариум - эоплейстоцен; в) лицевой отдел – эоплейстоцен – верхний плейстоцен;
г) нижняя челюсть – эоплейстоцен – верхний плейстоцен. Т.е. филогенетические
связи ископаемых гоминид Африки, Азии и Европы охватывают широкий диапазон:
эоплейстоцен – верхний плейстоцен. Заключая, максимальный объем случаев
трансгрессии величин краниометрических признаков ископаемых гоминид Азии и
Европы объясняется принадлежностью к одной части света – Евразии. Моногенизм
человеческого рода также обусловил значительную частоту случаев трансгрессии
ископаемых гоминид трех частей света. Филогенетические связи ископаемых гоминид
Азии и Европы охватывают широкий диапазон времени – чаще нижний и верхний плейстоцен.
Трансгрессия измерительных признаков черепа чаще всего видна для мозгового и
нижнечелюстного отделов, менее часто – для лицевого и, наконец, для
кальвариума.
Ключевые слова: антропология (физическая
антропология), палеоантропология, гоминиды Африки, гоминиды Азии, дивергенция
признаков
Вопрос о
дивергенции/трансгрессии краниометрических признаков плейстоценовых гоминид
Азии и Европы связан с многими актуальными проблемами современной палеоантропологии,
а, именно, просом о первоначального заселения Европы, судьбой азиатского очага
гоминизации, евразийскими миграциями гоминид и т.д. Сходство и различие ИГ Азии
и Европы связано с миграциями гоминид плейстоцена. Афроазиатские и афроевропейские
взаимоотношения гоминид описаны нами по литературным источникам в других
статьях.
Глубинное морфологическое
сходство ИГ Азии и Европы, в первую очередь, связано с прародиной человечества
Африкой и общей эволюцией гоминид до уровня эректусов и гейдельбержцев на Африканском
континенте. В аспекте нашей темы, сходство гоминид Азии и Африки обусловлено заселением
этих частей Евразии гоминидами одного эволюционного уровня. Начнем с самого
нижнего.
1. а) По мнению
А.П.Деревянко [2009], эректусы были широко распространены в Африке, Азии и,
вероятно, в Европе, они существовали в течение около 1,5 млн лет.
Он на основе археологических
свидетельств допускает возможность заселения Европы более 1 млн. лет назад.
Вопрос заключается в том, какова видовая принадлежность первых европейских
насельников?
В период 1-1,5 млн. лет
назад архантропы заселили южные районы Европы и Азии, вплоть до Индонезии.В
Австралию и Америку они не проникали. Эти архантропы представляют собой
наиболее массивных представителей древнейших людей и относятся к виду Homo
erectus (Дробышевский, 2009).
Указанный автор делит стадию архантропов на три временных отрезка:
нижнего эоплейстоцена 1,2-1, 8 млн лет), верхнего эуплейстоцена(800-1200
тыс.лет) и нижнего неоплейстоцена (400-800 тыс.лет). Останки архантропов
нижнего эоплейстоцена известны из Южной и Восточной Африки, а также, видимо, из
Грузии. На границе нижнего и верхнего эоплейстоцена или раньше архантропы
начали расселяться из Африки по тропической зоне Евразии. К началу
неоплейстоцена медленная экспансия происходила на север. В Европе население
рассматриваемого периода немногочисленно и концнтрировано в южных областях
(Дробышевский, 2004).
Вопрос об эволюционных
связях гоминид Азии и Европы встает, когда обсуждаются древнейшие гоминиды
Европы. Они открыты в местонахождении Атапуэрке (Испания), они размещаются в
диапазоне 1,2-0,5 млн. лет. Ископаемые останки гоминид, открытые в Гран-Долине,
древностью в 800 тыс.лет, позволили предположить существование на юге Европы
нового вида H.antecessor. Авторы этого предположения считают, что он имеет
аф-риканские корни, связан эволюционными корнями с эргасторами, и является
предком и неандертальцев Европы, и сапиенса. Таким образом, гейдельбержцы
Африки и Европы выводятся из родословной сапиенса. Первые представители H.
antecessor в среднем плей-стоцене мигрировали в Европу и дали начало
европейским неандертальцам. Африканские представители этого вида стали
предковыми для промежуточных видов – предков H.sapiens. В соответствии с другой
точкой зрения H. аntecessor мог быть потомком дма-нисцев, если допустить возможность
видообразования в Европе. Мы помним об африкан-ских корнях последних и
отнесении их к архантропам. Мигранты второй волны – архан-тропы могли вытеснить
или ассимировать H. аntecessor. Наконец, есть еще одна точка зрения: H.
аntecessor является результатом кладогенеза, произошедшего после ашельской миграционной волны [Деревянко,2009].
Указанный автор предположил,
что из Африки в Евразию и обратно в раннем палеолите было немного. Можно проследить
только два глобальных миграционных процесса. Каждая такая миграция
детерминировалась глубокими причинами(Деревянко,2009). Расселение человека в Европе началось в то
же время, что и в Азии. Это был единый процесс миграции из Африканской колыбели
человечества. Но архантропы быстрее всего заселяли близкие им по экологическим
условиям территории южной части Азиатского континента и в течение сравнительно
короткого времени вышли к Тихому океану. Значительно медленнее они могли
продвигаться на север в Европу через Ближний Восток, Малую Азию, Балканы и т.д.
или же через Северный Кавказ. Наиболее короткий путь человека из Африки в
Европу мог быть по сухопутному мосту. Но когда Европа соединилась с Африкой
или, по крайней мере, существовали мелководные, т.е. преодолимые для
архантропа, проливы при понижении уровня Мирового океана - эти проблемы требуют
своего разрешения.
А. П. Деревянко [2009]
обращает внимание на то, что многие антропологи отмечают значительную
эректоидную примесь у древнего населения Европы. Об этом свидетельст-вует
черепная коробка из Чепрано (Италия). Поэтому отрицание роли Homo erectus в
эво-люционном процессе в Европе неоправданно.
б) Вторая глобальная
миграция человека из Африки (с Ближнего Востока?) в Европу произошла около 600
тыс. л.н. Очень вероятно, что вторая
миграционная волна из Африки в Европу была связана с представителями Homo
heidelbergensis, для которых характерно сочетание неандерталоидных и сапиентных
черт.
В связи со второй
миграционной волной встает вопрос о судьбе автохтонного населения Европы. На
одних местонахождениях среднего плейстоцена преобладает ашельская индустрия, на
других прослеживается бифасиальная и галечная, на третьих отсутствуют рубила.
Видимо, с приходом новой миграционной волны в Европе происходил процесс не
замещения, а аккультурации. В этом отношении очень важны находки на
местонахожде-нии Сима-де-лос-Хуэзос (Испания) древностью более 530 тыс. лет.
Антропологические исследования около 4 тыс. человеческих костей показали, что
эти архантропы существен-но отличались от тех, чьи останки найдены в горизонте
ТД6 Гран-Долины, и были близки к неандертальцам. Палеоантропологические
материалы местонахождения Сима-де-лос-Хуэзос доказывают, что Европа была очагом
местной эволюционирующей ветви, впо-следствии давшей начало « классическому»
неандертальцу. Многие вопросы, связанные с двумя волнами миграции человека из
Африки в Европу, остаются дискуссионными [Деревянко,2009].
С. В. Дробышевский (2004)
пишет, что гоминиды времени 400-130 тыс.лет назад имели облась распространения
мало отличную от области распространения архантропов. Новые территории были
освоены только в северном направлении в Европе и, вероятно, в Азии.
Морфологическое разнообразие находок, датированных этим временем, весьма велико.
В промежутке времени
примерно от 500-400 до 130 тыс. лет назад на Земле обитало крайне разнообразное
население, имеющее множество переходных черт – примитивных и более
прогрессивных. Разные части света были населены людьми, сильно отли-чающимися
друг от друга. Часто гоминиды из древнейшей половины этого интервала
рассматриваются как Homo erectus, представители второй половины – вместе с
палеоантропами. Их особый промежуточный облик
позволяет рассматривать гоминид этого пласта вместе под обобщающим
названием Homo heidelbergensis.
Кроме прочего, существует
факт явной неоднородности уровня продвинутости синхронных гоминид в разных
регионах. В Юго-Восточной Азии в период 130-400 тыс. лет назад часты находки, определяемые как Homo
erectus; в Китае есть находки, определяемых как "Архаический Homo sapiens",
хотя их признаки далеко не так сапиентны, как у синхронных им африканских гоминид.
В Европе преобладали, видимо, Homo
heidelbergensis. В Африке в то же время появляются значительно более сапиентные
люди, иногда классифицируемые как Homo helmei, причём их датировки удревняются
даже до 490 тыс. лет назад (чаще даты для Homo helmei занимают промежуток от
100 до 200 тыс. лет назад). Примерно со 130-160 тыс. лет назад или даже ранее в
Африке и на Ближнем Востоке появляются представители вида Homo sapiens (хотя в
их организации немало архаичных признаков) (Дробышевский, 2009).
в) Палеоантропы, как и их
предки, продолжали мигрировать по планете, впрочем, расселение шло в основном к
северу по уже освоенным материкам; в Австралию и Америку они не проникали
(Дробышевский, 2009).
С. В. Дробышевский (2004) определяет время возникновения палеоантропов
в 150-100 тыс.лет, а исчезновения около 30 тыс.лет назад. Ареал распространения
их, связанный часто с миграциями: Европа, Западная Азия. Предполагаются
миграции неандертальцев в связи с климатическими причинами из Европы на Ближний
Восток, а также на восток до Ирака и Средней Азии. Такие миграции могли
начаться около 120 тыс. лет назад через Малую Азию, а далее в Центральную Азию
и Юго-западную Сибирь.
2. Далее сходство ископаемых
гоминид Азии и Европы может быть обусловлено широтными миграциями их между указанными
частями континента.
А. А. Зубовым (1995) рисует картину дифференциации человечества на
разных этапах протоистории на
одонтологических материалах. Уже у хабилисов
отмечен «западный» тип морфологии верхних резцов (нет лопатообразной формы), свойственный современным европеоидам и
негроидам Африки. Он характеризует и эректусов Африки, и некоторые ближневосточные
формы (Амуд, Кафзех), а также известные кроманьонские формы Европы (Сунгирь). В
Азии, начиная с древнейших эректусов, преобладает лопатообразная форма резцов
(«восточный» морфотип). Примечательно, что она встречается у европейских неандертальцев
и у некоторых ближневосточных гоминид (Схул, Табун), но в западных регионах не
сохраняется позже палеолита. Автор заметил, что у современных монголоидов есть
одонтоглифические особенности резцов, сближающие их с «западным» типом. А. А. Зубов
делает вывод, что независимого развития западного и восточного одонтологических
стволов не было, за исключением изолированных отрезков времени. Современный
восточный ствол формировался на основе западного за счет примеси местных
азиатских форм, характеризовавшихся классическим типом верхних резцов, который
мы видим у азиатских эректусов. Где и как происходила метисация – в Азии или в
Европе? В западном регионе, как и в восточном, повсеместно происходили
метисационные процессы, приводившие к появлению «синтетических» морфологических
вариантов зубов. Но носителей западного варианта на западе было больше, и при
метисации неандертальцев и кроманьонцев вклад первых (носителей «восточного»
варианта) был заметно менее половины. На востоке носителей «восточного» первоначального
варианта было больше, и в последующих поколениях он получает перевес.
Генетико-автоматические процессы в периоды изоляции надрасовых стволов привели
к концентрации носителей «западного» типа на западе и типа «восточного»
(обновленного за счет метисации) на востоке.
А. А. Зубов [1995] пишет еще
об одной особенности зубов. Имеется в виду гребень эпикристид, соединяющий
протоконид с метаконидом в середине тригонида. Данный признак отчетливо выражен
на зубах некоторых европейских неандертальцев, типичен для гейдельбержцев,
встречается у африканских архантропов и австралопитековых. Эпикристид маркирует
афро-европейскую ветвь эволюции ранних гоминид. Он редок на востоке и в современном
человечестве (при этом чаще всего встречается у негроидов и южных европеоидов).
Смешение сапиенса с европейскими неандертальцами не привело к увеличению
концентрации данного признака в Европе, так как число взаимодействующих неандертальских
групп было невелико. Далее, эта особенность одонтоглифики сочетается у европейских
неандертальцев с восточным типом устройства верхних резцов. В этом А. А. Зубов
видит сочетание западных и восточных особенностей у среднеевропейских гоминид,
следовательно, настаивает на «сетевидном» характере эволюции рода Homo.
А. А. Зубов (2004) писал,
что начальный этап дифференциации Homo sapiens
не был единомоментным, он охватывал период от раннего верхнего палеолита до
неолита (включительно), а, если учесть вклад периферийных форм – процесс уходит
корнями в средний палеолит. Важно, что вклад «восточного центра» в формирование
современного населения отвергать нельзя, хотя линия его собственной эволюции
угасла в конце среднего плейстоцена. Она была восстановлена за счет мигрантов с
запада представителей «позднего архаичного сапиенса». Позже новая миграция на
восток сформировавшегося сапиенса (Мальта и алтайские находки) привели к метисации
с местными группами и приняла участие в формировании монголоидного ствола
(Чжоукоудянь сходен с Сунгирем). «Сетевидная» эволюция ответственна за
появление разных «евро-монголоидных» и «монголо-австралоидных» вариантов.
Маркина Гора, Гримальди (на западе) и Лючан (на востоке) указывают на наличие периода,
когда Homo был разделен на два надрасовых конгломерата – «западный» и
«восточный» без четкой границы.
Доказательством этого
является следующее: реликтовые одонтоглифические признаки объединяют европейцев
и негроидов, австралийцев с монголоидами. По данным генетики первое деление
внутри вида имеет древность 92 тыс.лет, деление внутри больших стволов – 39 тыс.лет.
Бушмены и австралийцы выделились первыми- 60-90 тыс. лет, а сохранились
благодаря изоляции. В направление к нашим дням происходит изживание архаики в
морфологии людей. Чаще всего, идентификация субтаксонов возможно в мезолите -
(неолите) – верхнем палеолите. В первичном «восточном стволе» сформировались
монголоидный и австралоидный компоненты, а «западный» ствол дал европеоидный и
негроидный компоненты сапиенса. Процесс дифференциации имеет непрерывный
характер.
Дивергенция расовых стволов
происходила, по данным генетики, в пределах последних 100 тыс.лет. Ранее всех
выделилась ветвь монголоидов (92 тыс.лет), протонегроиды и протоевропейцы
разделились 33 тыс.лет назад. В Африке формирование экваториальных комплексов
происходило еще раньше. Монголоидный и
негроидный комплексы оформились окончательно позже. Европеодная и монгоидная
расы наиболее метисированы.
Материал и
метод исследования
В нашей работе сделана
попытка выявить пространственную структуру краниометри-ческих признаков
гоминид, охватывающую три части света (Африку, Азию, Европу) с помощью
результатов канонического анализа, проведенного на основе системы
HomoBase-Kanoklass, созданной профессором Е. В. Дерябина. Канонический анализ
объективно отра-жает сходство и различие ансамблей краниометрических признаков
ископаемых гоминид.
Материалом для работы
послужили данные по краниометрии ископаемых гоминид плей-стоцена Африки, Азии
Европы, оформленные в виде РС базы данных , разработанной д.б.н. В. Е. Дерябиным.
Указанная база включает измерения черепа 250 гоминид. В на-стоящей работе
обсуждены результаты канонического анализа 82 ископаемых гоминид трех частей
света. Имеются в виду: а) австралопитеки – 3 вида; б) H. habilis, в) H.
rudolf-ensis, г) H. ergaster, д) H. erecrus, е) H. heidelbergensis, ж) H. neanderthalensis,
з) архаичный H. sapiens.
В работе проанализированы 191 график канонического анализа краниометрических
признаков ископаемых гоминид и их предшественников (австралопитековых). Первоначальный
анализ показал, что ситуации пространственного взаимного положения группировок
ископаемых гоминид на материале краниометрических признаков были распределены
по возрастающей между следующими вариантами:
а) Трансгрессия
гоминид Европы и Азии; Африка отдельна; 33, 94% (93 случая),
б) Трансгрессия гоминид
Африки, Азии и Европы; 29,92% (82 случая),
в) Трансгрессия гоминид
Африки и Азии; Европа отдельно; 14,62% (40 случаев),
г) Трансгрессия гоминид
Африки и Европы; Азия отдельно. 11,31% (31 случай),
д) Отсутствие трансгрессии
гоминид Африки, Европы и Азии; 10,22%(28 случаев).
Итого-274 случая. Итоговая
величина числа случаев, которая превышает количество графиков канонического
анализа, объясняется тем, что в случаи отсутствия гоминид одной части света, мы
присоединяли данный вариант ко всем возможным.
Анализ всей изученной
совокупности случаев трансгрессии географических групп гоминид позволяет
заключить следующее. Объем связей гоминид Азии и Европы в 63,86%. Совокупные
связи гоминид Африки и Азии могут быть оценены в 44,54%, они же у гоминид
Африки и Европы в 40,69%. В настоящей работе конкретно обсуждается вопрос филогенетического
взаимоотношения плейстоценовых гоминид Азии
и Европы на материале краниометрических признаков. В совокупности они
представлены вариантами «а» и «б».
Явления трансгрессии и
дивергенции признаков черепа гоминид были выявлены с помощью канонического анализа диаметров следующих
отделов черепа. В частности:
1.Европа+Азия; Африка
отдельно (МО=43; МО+ЛО=3; ЛО=21; НЧ=26)
2.Африка+Азия+Европа (МО=38;
МО+ЛО=5; ЛО=14; НЧ=25)
Мы видим, что в первом и
втором случаи (наиболее часто встречаемом) повышена частота анализов нижней
челюсти, можно говорить также о повышенной частоте встречаемости анализов
признаков мозгового и лицевого отделов черепа.
Вариант – Азия+ Европа;
Африка отдельно.
При первоначальном анализе
данный вариант пространственного взаиморасположения географических группировок
ископаемых гоминид был поддержан следующими отделами черепа (см. выше). В нашем
уточненном анализе – МО=46; МО+ЛО=4; ЛО=18; НЧ=24. Различие почти не ощутимое.
Соотношение отделов черепа, дающих указанное разбиение ископаемых гоминид почти
одинаковое.
Далее проанализируем отделы
черепа по признаку видовой принадлежности ископаемых гоминид, показывающих данное
разбиение.
Мозговой отдел.
а) Количество случаев варианта «Азия+Европа; Африка
отдельно» - 39. Они проявились при анализах: эректусы- 21; гейдельбержцы- 2;
неандертальцы- 10; архаичный сапиенс- 6 (рис.1-24). На рис.1 мы видим трансгрессию
значений признаков мозгового отдела гоминид Азии (от эректусы, архаичные сапиенсы, кроманьонцев) и Европы (неандертальцы,
сапиенсы) и отдельное расположение кластера африканских гоминид (гейдельбержцы).
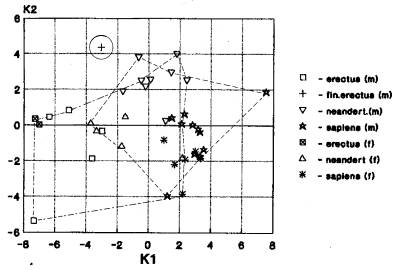
Указанные гоминиды
располагаются в диапазоне «нижний плейстоцен - верхний плейстоцен». Можно
считать, что филогенетические связи гоминид Азии и Европы во времени были
значительными и постоянными.
б) Среди анализов выявлен вариант, в котором наблюдается совмещение кластеров Азии и Европы, но при этом африканский кластер сопряжен или с Азией (б-1)(рис.2а-39), или Европой (б-2)(рис.2б-191). На рис. 2а мы видим трансгрессию азиатского кластера (питекантропы, неандертальцы,кроманьонцы) и европейского (гейдельбержцы, неандертальцы, кроманьонцы) и трансгрессию азиатского кластера и африканского (гейдельбержцы).
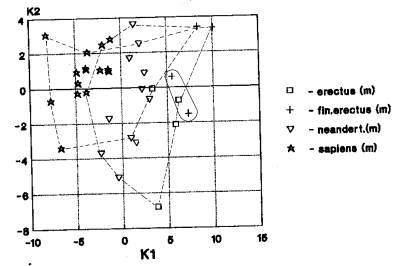
Подвариант б-1 проявился: у
рудольфензис- 2; эректусов-1; гейдельбержцев-1; неандертальцев-1. Данные
гоминиды размещены в диапазоне времени от эоплейстоцена до верхнего
плейстоцена.
Подвариант б-2 проявился у
неандертальцев (2 случая) (верхний плейстоцен). Итого два подвариант «б»
представлены в 7 случаях. Верхнеплейстоценовый вариант «б» менее многочислен
чем вариант «а». Несколько выделяется H.rudolfensis в варианте расположения
«Азия (Африка) + Европа», что, видимо, обусловлено очень ранними филогенетическими
связями гоминид Африки и Азии.
Кальвариум. Общее количество
случаев-4.
а) Количество случаев варианта «Азия+Европа; Африка отдельно»
-3. Он проявился при анализах гоминид эоплейстоцена, нижнего и среднего плейстоцена
(рис.3-6). На рис. 6 мы видим совмещение кластеров гоминид Азии (архаичные
сапиенсы, кроманьонцы) и Европы (неандертальцы и кроманьонцы), а также
раздельное положение африканского кластера (эректусы и гейдельбержцы).
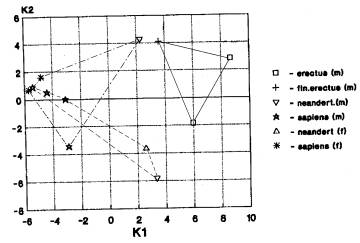
б) В одном случаи при
анализе раннего Homo проявился вариант «Азия (Африка) +Европа», он датируется
эоплейстоценом и отражает древние связи гоминид Африки и Азии.
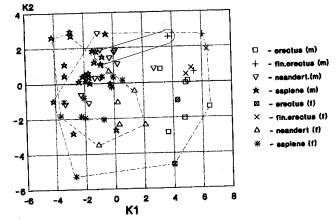
Лицевой отдел. Количество
случаев-18.
а) Вариант «Азия+Европа; Африка отдельно» проявился при анализе: гейдельбержцы-4; неандертальцы-10. (рис.4-85). На рисунке 4 мы видим пространственное совмещение азиатского кластера (неандертальцы и кроманьонцы) и европейского (неандертальцы и кроманьонцы), а также изолированное положение африканского кластера (эректусы и гейдельбержцы).Таким образом, подобное разбиение географических кластеров ископаемых гоминид встречается в среднем плейстоцене и верхнем плейстоцене, главным образом.
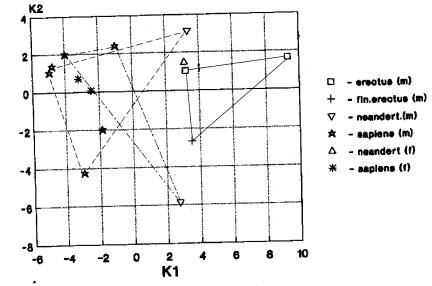
б) Вариант «Азия (Африка)
+Европа» проявляется при анализах неандертальцев (1) и архаичного сапиенса (1).
Речь идет здесь о верхнеплейстоценовых гоминидах. Таким образом, признаки
лицевого отдела свидетельствуют об эволюционных связях ископаемых гоминид Азии
и Европы в более поздний период чем признаки мозгового отдела и кальвариума.
Нижняя челюсть. Количество
случаев-24.
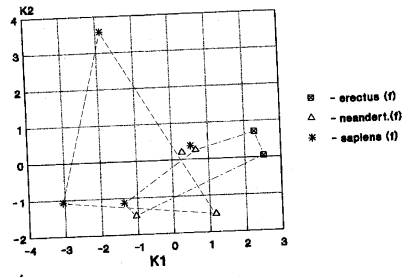
а) Вариант «Азия+Европа; Африка отдельно» проявился при анализе: эректусов-2; гейдельбержцев-2; неандертальцев-13 (рис.5-171). На рис.5 мы видим пространственное совмещение кластеров азиатских гоминид (неандертальцы и кроманьонцы) и европейских (неандертальцы и кроманьонцы) при изолированном от них расположении африканских питекантропов и кроманьонцев. Указанные гоминиды размещаются в нижнем, среднем и верхнем плейстоцене.
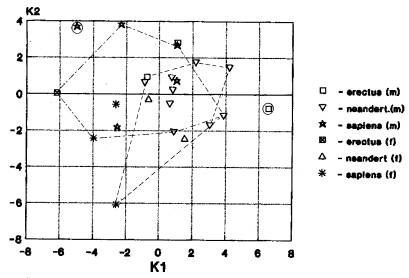
б) Вариант «Азия+Европа» выявлен в 7 случаях: эректусы-2; неандертальцы-4; архаичные сапиенсы -1. (рис.6-95). На рис.6 мы видим пространственное совмещение кластеров азиатских гоминид (эректусы и неандертальцы) и европейских (неандертальцы и кроманьонцы). Указанные гоминиды обозначают хронологический диапазон от нижнего до верхнего плейстоцена.
Таким образом, мы видим что
ископаемые гоминиды Азии и Европы демонстрируют трансгрессию признаков в следующих
диапазонах времени:
а) Мозговой отдел –
нижний-верхний плейстоцен или эоплейстоцен-верхний плейстоцен.
б) Кальвариум -
эоплейстоцен-средний плейстоцен.
в) Лицевой отдел - средний-верхний плейстоцен,
г) Нижняя челюсть-
нижний-верхний плейстоцен,
Т.е. филогенетические связи
ископаемых гоминид Азии и Европы охватывают широкий диапазон –
эоплейстоцен-верхний плейстоцен, но чаще нижний-верхний плейстоцен.
Трансгрессия
краниометрических признаков ископаемых гоминид Азии и Европы осуществляется за
счет мозгового отдела, реже - нижней челюсти, лицевого отдела и кальвариума. Чаще
всего за счет первых трех отделов черепа.
Вариант- Африка+Азия+Европа
Данный вариант взаимного
расположения географических группировок ископаемых гоминид трех частей света
второй по величине среди рассмотренных вариантов. При первоначальном анализе (с
учетом всех возможных ньюансов) он поддержан следующими отделами черепа:
МО=38; МО=Ло=5; ЛО=14; НЧ=25.
В уточненном (строгом) виде
видно, что вариант с трансгрессией значений измерительных признаков черепа
ископаемых гоминид трех частей света проявился:
МО=15; МО+ЛО= 5; ЛО=10; НЧ=21,
т.е. соотношение отделов черепа близко в обоих
вариантах. Мы видим, что трансгрессия измерительных признаков черепа ископаемых
гоминид Азии и Европы осуществляется, в первую очередь, за счет нижней челюсти
и мозгового отдела, далее за счет лицевого отдела и, наконец, за счет кальвариума.
Практически все традиционные отделы принимают участие в явлении трансгрессии
признаков.
Мозговой отдел. Количество
случаев-15.
Вариант пространственного
расположения географических групп гоминид «Африка+Азия+Европа» проявляется при
анализе следующих гоминид: гейдельбержцы-8; неандертальцы-3, эректусы-2,
архаичные сапиенсы-2 (рис.7-42). На рис.7 видна трансгрессия ареалов значений
гоминид Африки, Азии и Европы на материале признаков мозгового отдела. Африканский
кластер представлен гейдельбержцами и кроманьонцами, азиатский – эректусами,
гейдельбержцами, архаичными сапиенсами и кроманьонцами, европейский кластер –
неандертальцами и кроманьонцами. Таким образом, для признаков мозгового отдела
вариант трансгрессии признаков мозгового отдела существовал с нижнего до верхнего
плейстоцена.
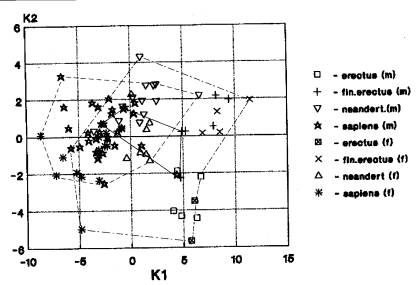
Кальвариум. Количество
случаев-5.
Описываемый вариант пространственного расположения географических групп гоминид наблюдается на материале измерительных признаков при анализах: австралопитеки-4; хабалисы-1 (рис.8-17). Признаки кальвариума позволяют наблюдать трансгрессию кластеров Африки, Азии и Европы. Африканский кластер включает грацильных и массивных австралопитеков, азиатский – кроманьонцев, а европейский – неандертальцев и кроманьонцев. Анализы они приурочены к эоплейстоцену.
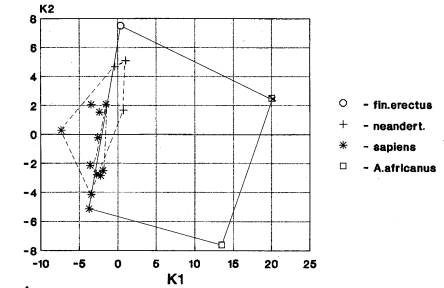
Лицевой отдел. Количество
случаев-10.
Анализируемый вариант пространственного расположения географических групп гоминид наблюдается на материале краниометрических признаков лицевого отдела при анализах: неандертальцы-5; рудольфензис-2; эректусы-2, гейдельбержцы-1 (рис.9-143). На рис. 8 видна одновременная трансгрессия признаков лицевого отдела у гоминид Африки, Азии и Европы. Африканский кластер образован неандертальцами, гейдельбержцами и кроманьонцами, азиатский – неандертальцами, архаичными сапиенсами и кроманьонцами, европейский – гейдельбержцами, неандертальцами и кроманьонцами. Таким образом, здесь лидируют гоминиды верхнего плейстоцена, далее следуют гоминиды среднего и нижнего плейстоцена, эоплейстоцена.
Нижняя челюсть. Количество
случаев-21.
Исследуемый вариант взаимного пространственного расположения географических групп гоминид наблюдается на материале измерительных признаков нижней челюсти при анализах: эректусы и неандертальцы- 7 и 7; архаичные сапиенсы- 3; хабилисы и гейдельбержцы – 2 и 2 (рис. 10-20). На рис. 20 видна трансгрессия нижнечелюстных признаков гоминид Африки, Азии и Европы. Африканский кластер образован хабилисами, эректусами и кроманьонцами, азиатский – эректусами, неандертальцами, архаичными сапиенсами и кроманьонцами. Европейский кластер включает неандертальцев и кроманьонцев. Т.е. чаще всего трансгрессия признаков у гоминид трех частей света проявляется в нижнем и верхнем плейстоцене. А вообще диапазон времени проявления трансгрессии широк - от эоплейстоцена до верхнего плейстоцена.
Суммируя все написанное о
случаях трансгрессии краниометрических признаков ископаемых гоминид трех частей
света – Африки, Азии и Европы, можно представить за счет каких отделов черепа и
в какое время она осуществлялась:
А) Мозговой отдел –
нижний-верхний плейстоцен;
Б) Кальвариум -
эоплейстоцен;
В) Лицевой отдел –
эоплейстоцен – верхний плейстоцен;
Г) Нижняя челюсть –
эоплейстоцен – верхний плейстоцен.
Мы видим, что
филогенетические связи ископаемых гоминид Африки, Азии и Европы охватывают
широкий диапазон: эоплейстоцен – верхний плейстоцен.
Заключение
1. Максимальный объем случаев трансгрессии величин краниометрических признаков ископаемых гоминид Азии и Европы достаточно легко объясняется: речь идет о связях гоминид одной части света – Евразии. Моногенизм человеческого рода также обусловил значительную частоту случаев трансгрессии ископаемых гоминид трех частей света.
2.
Филогенетические
связи ископаемых гоминид Азии и Европы охватывают широкий диапазон времени –
чаще нижний и верхний плейстоцен.
3.
Трансгрессия
измерительных признаков черепа чаще всего видна для мозгового и нижнечелюстного
отделов, менее часто – для лицевого, и еще менее часто для кальвариума.
Литература
1.
Зубов
А.А. Проблемы внутриродовой систематики рода НОМО в связи с современными представлениями
о биологической дифференциации человечества//Современная антропология и
генетика и проблема рас у человека. М.: Институт этнологии и антропологии РАН,
1995. С.18-43.
2.
Зубов
А.А. Палеоантропологическая родословная человека. М.: Институт этнологии и антропологии
РАН, 2004, С.552.
3.
Деревянко
А.П. Древнейшие миграции человека в Евразии в раннем палеолите. Новосибирск,
2009, С.231.
4. Дерябин
В.Е., Харитонов В.М. Объективная классификация рода Homo на основе многомерного биометрического анализа
признаков черепа гоминид//Вопросы антропологии, вып.91. М.,2003. С.22-34.
5.
Дробышевский
С.В. Предшественники. Предки? Часть III. Архантропы. Часть IY: Гоминиды,
переходные от архантропов к палеоантропам.- 2004. 344 с.
6.
Дробышевский
С.В. «Антропогенез». Учебное пособие для вузов. М., 2009, С.45
7.
Рогинский
Я.Я. Проблемы антропогенеза. М.: Изд-во МГУ,1969, С.262.
SIGNS OF THE SKULL DIVERGENCE OF FOSSIL HOMINIDS
IN ASIA AND EUROPE
Kharitonov V. M.
We investigated the divergence (transgression) of craniometric characters
in allopatric speciation of hominids. Analyzed about 190 plots of canonical
analysis of cranial size of fossil hominids. Material are the craniometric
features 250 fossil hominids Pleistocene of Africa, Asia and Europe. Variants
of the relative position of the hominid geographic groups: 1) Europe + Asia,
Africa separately - 33.94% 2) Africa +
Asia + Europe - 29.92% 3) Africa + Asia, Europe, but individual -14.62 % 4)
Africa + Europe, Asia-11.31% separately. 4) Africa alone, separate but Asia,
Europe alone - 10.22%. Cumulative connection of hominids in Africa and Asia -
44.54%, the second-minid Africa and Europe - 40.69%, and hominids in Asia and
Europe - 63.86%. Divergence (transgression)
hominid traits were due to different degrees by different departments of the
skull. Identified the following options for the relative position of
geographical clusters of hominids. Option - Asia + Europe, Africa alone. Fossil
hominids in Asia and Europe show signs of transgression in the following ranges
of time: a) cerebral Department - Lower-Upper Pleistocene or upper
Pleistocene-Eopleistocene b) kalvarium-Eopleistocene-middle Pleistocene. c) the
front office - middle-upper Pleistocene, d) lower jaw- lower-upper Pleistocene.
Phylogenetic relationships used dig hominids in Asia and Europe cover a wide
range - Eopleistocene-Upper Pleistocene, but most often lower - upper
Pleistocene. Transgression craniometrical features of fossil hominids in Asia
and Europe at the expense of the brain, at least - the lower jaw, facial and
kalvariuma department. Most often at the expense of first three sections of
skull. Option-Africa+ Asia+ Europe. Transgression of craniometric traits was carried out: a) cerebral Department - Lower-Upper Pleistocene, and b) kalvarium-Eopleistocene
c) the front office - Eopleistocene - Upper Pleistocene, and d) lower jaw -
Eopleistocene - the upper Pleistocene. I.e. phylogenetic relationships of
fossil hominids from Africa, Asia and Europe cover a wide range Eopleistocene -
upper Pleistocene. In summary the maximum transgression of the cases the values
of craniometric traits of
fossil hominids in Asia and Europe due to belonging to one of the world -
Eurasia. Monogenizm human species also caused a significant incidence of transgression
of the fossil hominids of the three parts of the world. Phylogenetic relationships
of fossil hominids in Asia and Europe cover a wide range of time - Lower and Upper Pleistocene. Transgression
of measuring the skull traits most often visible to the brain and the
mandibular divisions, often less for
front office and, at last, for kaivarium.
Key words: anthropology (physical anthropology),
paleoanthropology, hominid African, hominids in Asia, signs of divergence.
НИИ и Музей антропологии МГУ, Москва
Institute and Museum of Anthropology, MSU, Moscow
Поступила в редакцию
19.06.2012.