Математическая
морфология.
Электронный
математический и медико-биологический журнал. - Т. 14. -
Вып. 1. - 2015. -
URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-45-html/TITL-45.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-45-html/cont.htm
УДК 551
Лекция
ДНК-генеалогия как молекулярная история
Ó 2015 г. Клёсов А. А.
Изложены основные понятия и
определения новой научной дисциплины, формирующейся в последние несколько лет –
ДНК-генеалогии, и показаны ее принципиальные отличия от популяционной генетики
человека. Методологической базой новой дисциплины является использование
аппарата химической и биологической кинетики для анализа картины
неупорядоченных мутаций в определенных нерекомбинантных участках Y-хромосомы
ДНК. Цель анализа – перевод динамической картины мутаций в хронологические,
временные показатели, датировки времен жизни общих предков популяций, древних
родов и племен. Как иллюстрации, приведены примеры определения
времен жизни древних общих предков этнических киргизов гаплогруппы R1a,
а также современных немцев и индийцев той же гаплогруппы. Общие
предки двух групп современных киргизов рода R1a (самый представительный род,
или гаплогруппа в Киргизии) – по «записям» в их Y-хромосоме – жили 1325±290 и
775±450 лет назад (первая группа является в Киргизии преобладающей по данным
ограниченных выборок), общий предок этих двух групп жил примерно 1650 лет
назад, то есть примерно в 4-м веке нашей эры. Это, наиболее вероятно, скифы
или сарматы, хотя здесь ДНК-генеалогия должна согласовать свои предположения с
исторической наукой, и либо ее дополнить, либо, напротив, откорректировать свои
предположения. Общий предок современных немцев гаплогруппы R1a жил – по разным
выборкам 2008 и 2014 гг и по результатом расчетов по
гаплотипам разной протяженности - 4800±400, 4300±300, 4575±560 и 4675±510 лет
назад, что показывает приемлемую репрезентативность выборок и диапазон погрешностей
расчетов в ДНК-генеалогии при параллельном использовании 12-, 25-, 67-, и
111-маркерных гаплотипов. Этот вопрос является ключевым в ДНК-генеалогии, а
именно как можно по ограниченным выборкам в десятки и сотни гаплотипов делать
(пусть и предварительные) выводы о временах жизни общих предков популяций в
миллионы человек. В данном случае расчеты ДНК-генеалогии были подтверждены
археологическими раскопками, а именно датировками 4600 лет назад для ископаемых
носителей гаплогруппы R1a в Германии (Haak et al., 2008). Общий предок
современных индийцев гаплогруппы R1a, в значительной степени (до 72%)
составляющих высшие касты Индии, жил 4750±500 лет назад, что в пределах погрешности
расчетов совпадает со временем жизни общего предка этнических русских (4900±500
лет назад), и их предковые гаплотипы почти идентичны. Излагаемый подход
фактически закладывает основы «молекулярной истории», в которой функцию основного
инструментария выполняет высокотехнический анализ молекул ДНК современников и
ископаемых останков.
Ключевые слова: ДНК-генеалогия, гаплотип,
гаплогруппа, мутации, Y-хромосома.
Начнем с понятия
«молекулярная история». Суть его в том, что становится возможным
следить за передвижениями древних народов не с помощью лопаты и кисточки археолога,
не обмеряя черепа, не хитроумно расплетая созвучия и значения слов в живых и
мертвых языках, не изучая древние фолианты в библиотеках и монастырях, а просто
прослеживая за метками в Y-хромосомах наших ДНК. Они,
эти метки, не могут «ассимилироваться», или «поглотиться» другими языками,
культурами, или народами, как это происходит тысячелетиями с языками,
культурами, народами в рамках понятий истории, лингвистики, этнографии, антропологии.
Иначе говоря, методология новой исторической науки, «молекулярной истории», или
ДНК-генеалогии, основывается на изучении молекул нуклеиновых кислот, а именно
ДНК, дезоксирибонуклеиновой кислоты, в человеческих организмах, как живых, так
и в древних костных останках. То, что еще несколько лет назад казалось каким-то
развлечением, оказалось, дает истории, антропологии, археологии, лингвистике
возможность проверить концепцию, рассмотреть данные под принципиально другим
углом, связать воедино казалось бы разрозненные части
общей картины наших знаний об окружающем мире. Разумеется, не следует понимать
буквально слова выше, как то, что отныне не нужно читать фолианты, изучать
языки, и не проводить археологических раскопок. Напротив, это все приобретает
еще большую значимость и ценность, поскольку усиливает выводы традиционных
подходов в совокупности с выводами ДНК-генеалогии, при условии взаимного их согласования.
Или, напротив, заставляет пересматривать казалось бы
устоявшиеся выводы, если они входят в явное противоречие с данными
ДНК-генеалогии.
В этой cтатье
речь пойдет о новой науке, которая только создается, основы которой только
закладываются в последние годы. Если точнее, то основы этой
новой науки уже созданы, причем стремительно (Klyosov, 2008; Klyosov, 2009a,
2009b; 2009c; Клёсов, 2011; Rozhanskii, Klyosov, 2011, 2012; Klyosov, 2012;
Klyosov, Rozhanskii, 2012a, 2012b; Klyosov et al., 2012; Klyosov, Tomezzoli,
2013; Клёсов, 2013а, Клёсов, 2013b; Klyosov, Mironova, 2013; Elhaik et al.,
2014; Klyosov, 2014), и идет прогрессивное накопление экспериментального
материала. Новый материал поступает потоком, ежедневно в базы данных
уходят десятки и сотни новых «экспериментальных точек», которые по принципу
обратной связи корректируют методологию новой науки, что приводит к уточнению методов
расчета.
Имя этой науки —
ДНК-генеалогия. Ее экспериментальные данные — это картина мутаций в нерекомбинантных
(то есть не комбинирующихся с другими в ходе передачи наследственной информации
потомкам) участках мужской половой хромосомы (на самом деле — и в
митохондриальной ДНК, но в этой статье речь пойдет о Y-хромосоме, более
информативной для исторических исследований), причем картина мутаций
как в Y-хромосомах отдельных людей, так и их групп, племен, родов, популяций.
Естественно, в данном контексте термин «популяций» относится к мужской половине
рассматриваемой популяции. Методология новой науки — перевод динамической
картины мутаций в хронологические показатели, во времена жизни общих предков
популяций, а на самом деле — общих предков древних родов и племен. То есть фактически
по мутациям в ДНК производится расчет времен, когда в
древности жили эти рода и племена, и как эти времена меняются от территории к
территории, что может указывать на направления древних миграций (Klyosov,
2009a,2009b, 2009c; Klyosov, 2011; Rozhanskii, Klyosov, 2011; Klyosov, 2012).
О каких «экспериментальных
точках», или «экспериментальных данных» здесь идет речь? Что за «динамическая
картина мутаций» в Y-хромосоме? Как это – переводить «картину мутаций» в
хронологические показатели?
Результаты ДНК-тестирования
мужчин в данном случае сводятся к двум основным показателям. Это – гаплотип и
гаплогруппа. Гаплотип – это набор чисел, который является «личным номером
ДНК-паспорта» любого мужчины, без исключения. А гаплогруппа, фигурально говоря,
это обложка паспорта. Это – не индивидуальная, а групповая характеристика.
Гаплогруппа определяет носителя гаплотипа в определенный род, или племя. У
любой гаплогруппы, как у любого рода или племени, был один общий предок,
патриарх. Поэтому как номер паспорта при наличии обложки является уникальным «определителем»
человека, так и гаплотип в совокупности с гаплогруппой являются не менее
уникальным определителем человека, при условии, что номер (гаплотип) является
достаточно протяженным. Примеры дадим ниже.
Если у мужчины такого номера
и паспорта нет, то это не мужчина, а женщина. Именно к такому выводу сейчас
нередко приходят археологи, которые именно так наиболее надежно определяют пол
скелетного остатка в древнем захоронении. Нет Y-хромосомы, нет мужского гаплотипа и мужской гаплогруппы – значит, женщина.
У последних – свой ДНК-паспорт, под названием
«митохондриальная ДНК», который они передают своим детям, как девочкам, так и
мальчикам. А отец передает исключительно Y-хромосому, и только, разумеется,
сыновьям.
Таким образом
у каждого мужчины или мальчика есть Y-хромосома (которой, повторяю, у женщин
нет), которая с большой точностью передается к сыну от отца, а тому – от своего
отца, а тому – от своего, и так на протяжении сотен, тысяч, десятков и сотен тысяч
лет, и все эти тысячелетия, поколение за поколением, Y-хромосома передается
почти в неизменном виде. «Почти» - это потому что в мире ничего неизменного –
на протяжении тысячелетий - не бывает. В Y-хромосоме тоже происходят
постепенные (точнее, микроскачками) изменения, и в среднем на каждое поколение,
от каждого отца к каждому сыну, в Y-хромосоме происходит одна мутация, при
которой один нуклеотид (это – минимальный блок ДНК, размером примерно в
молекулу обычного столового сахара) случайно, неупорядоченно превращается в
другой. А в Y-хромосоме таких нуклеотидов 58 миллионов. Так что «почти в
неизменном виде» - это одна мутация на 58 миллионов потенциальных вариантов,
которые в остальном остаются неизменными. Каждая такая мутация наследуется
практически навсегда, потому что при копировании ДНК в следующем поколении
биологическая копировальная машина в клеточном ядре «не знает», как оно там
было в прошлом поколении. Ее задача – копировать.
Поэтому у каждого мужчины в Y-хромосоме своя картина
мутаций. У близких родственников картина мутаций похожа, потому что они,
родственники, сравнительно недавно произошли от одного общего предка, и
дополнительные мутации в их Y-хромосомах еще не успели образоваться. Если общий
предок жил тысячелетия назад, то картина мутаций у потомков разная. Как эту
картину отображают?
Отображают картину мутаций в виде так называемых гаплотипов, упомянутых выше как «личный паспорт» каждого мужчины, а именно в виде определенных последовательностей чисел, характерных для каждого человека. Чем эта последовательность длиннее, тем уникальнее гаплотип. Например, у автора этих строк гаплотип Y-хромосомы, состоящий из 111 чисел, записывается в следующем виде:
13 24 16 11 11 15 12 12 10 13 11 17 16 9 10 11 11 24 14 20 34 15 15 16 16 11 11 19 23 15
16 17 21 36 41 12 11 11 9 17 17 8 11 10 8 10 10 12 22 22 15 10 12 12 13 8 15 23 21 12 13
11 13 11 11 12 13 31 15 9 15 12 25 27 19 12 12 12 12 10 9 12 11 10 11 12 30 12 14 25 13
9 10 18 15 20 12 24 15 12 15 24 12 23 19 11 15 17 9 11 11
Это — так называемый 111-маркерный гаплотип. Здесь первое число показывает, что в одном из маркеров, или локусов Y-хромосомы определенная последовательность нуклеотидов повторяется 13 раз. В другом маркере, который здесь показан как следующий, блок нуклеотидов повторяется 24 раза, и так далее.
Покажем для сравнения 67-маркерный гаплотип киргиза Тазабекова, из Киргизского Проекта FTDNA, о котором речь пойдет ниже:
13 25 16 11 11 14 12 12 10 13 11 18 15 9 10 11 11 23 14 21 32 12 15 15 16 11 11 19 23 16
16 19 17 33 40 14 11 11 8 17 17 8 11 10 8 12 10 12 21 22 15 10 12 12 13 8 14 22 21 12 12
11 13 11 11 12 12
И еще один, киргиза Чороева:
13 26 16 11 11 11 12 12 10 14 11 17 15 9 10 11 11 24 14 21 31 12 15 15 16 11 11 19 23 16
16 18 17 33 40 15 11 11 8 17 17 8 11 10 8 12 10 12 21 22 15 10 12 12 13 8 13 22 21 12 12
11 13 11 11 12 12
Выделены мутации между моим гаплотипом и каждым из приведенных киргизских. Их, соответственно, 31 и 33 мутации, а между двумя киргизскими – всего 9 мутаций. Уже видно, что оба киргиза между собой намного более близкие родственники, чем каждый из них со мной. Здесь мутации в ряде маркеров подсчитывались специальным образом, но не будем сейчас на этом останавливаться. Эти различия можно перевести в хронологические показатели, зная среднюю скорость образования мутаций в гаплотипах, и получится, что общий предок автора этих слов и указанных киргизов жил примерно 4500 лет назад, в середине III тыс до н.э., как раз во времена арийских миграций по Русской равнине и Средней Азии, а обоих киргизов – примерно 1010 лет назад, в начале 11-го века нашей эры. Как делается такой расчет, мы покажем ниже.
Откуда получают те числа,
которые показаны в гаплотипах? Всё это на самом деле четко определено и
зафиксировано (Интернет-сайт «Sorenson Molecular Genealogy Foundation»). В первом маркере (под названием
DYS393) повторяется четверка нуклеотидов AGAT, то есть
аденин-гуанин-аденин-тимин
GTGGTCTTCTACTTGTGTCAATAC/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/AGAT/ATGTATGTCTTTTCTATGAGACATACCTCATTTTTTGGACTTGAGTTC
и эти повторы обрамляются уже неупорядоченными
последовательностями нуклеотидов в ДНК, как показано выше. Этот участок в ДНК и
есть локус, он же маркер DYS393. Во втором маркере, под названием DYS390,
повторяется уже другая четверка, TCTG, то есть тимин-цитозин-тимин-гуанин, которая
переходит в четверку TCTA, то есть тимин-цитозин-тимин-аденин, и число повторов
складывается:
TATATTTTACACATTTTTGGGCCCTGCATTTTGGTACCCCATAATATATTCTATCTA/TCTG/TCTG/TCTG/TCTG/TCTG/TCTG/TCTG/TCTG/TCTA/TCTA/TCTA/TCTA/TCTATCTA/TCTA/TCTA/TCTA/TCTA/TCTA/TCTA/TCTG/TCTA/TCTA/TCTA/TCATCTATCTATCTTTCCTTGTTTCTGAGTATACACATTGCAATGTTTTCATTTTACTGTCAC
Перечисленные четыре нуклеотида — это язык ДНК, который состоит из этих четырех слов, или четырех букв, при сокращенной записи. Эти нуклеотиды комбинируются парами, их так и называют — «пары оснований». Они фактически держат двойную спираль ДНК. Нам эти четверки (а также тройки нуклеотидов, и другие типы повторов) в маркерах ДНК в дальнейшем не понадобятся, это описание мы даем только для того, чтобы показать, что за числами в гаплотипах стоят масштабные исследования, которыми занимаются в мире сотни и тысячи человек.
Мы видим, что и у меня, и у указанных киргизов первый маркер дал одинаковое число повторов, 13. Одинаковое число повторов у всех троих и в третьем маркере (16), и в четвертом (11), и в пятом (11), седьмом (12), восьмом (12), девятом (10) и одиннадцатом (11) маркерах. Это не удивительно, потому что величины констант скоростей мутаций в большинстве этих стабильных маркеров одни из самых низких, и составляют в пересчете на 25 лет, то есть на условное поколение, 0.0005 (первый маркер), 0.00009 (седьмой маркер), 0.00022 (восьмой маркер), 0.0003 (одиннадцатый маркер). Это означает, что мутация в этих маркеров происходит в среднем раз в 50 тысяч лет, 280 тысяч лет, 115 тысяч лет, 85 тысяч лет, соответственно. Поэтому эти маркеры весьма стабильные. А вот второй маркер имеет константу скорости мутации 0.0027 на условное поколение, то есть маркер в среднем мутирует раз в 9 тысяч лет. Вот он и мутировал дважды при переходе от общего предка, который жил примерно 4500 лет назад, ко всем нам троим. Десятый по счету маркер еще более «быстрый», с константой скорости мутации 0.0043 на условное поколение, мутирует в среднем раз в 6 тысяч лет, вот он и мутировал у одного из нас троих. Но единичные мутации не дают хорошей статистики, поэтому желательно рассматривать большие выборки гаплотипов, в сотни, а то и в тысячи гаплотипов. В среднем же 67-маркерный гаплотип мутирует с константой скорости мутации 0.12 за 25 лет, то есть одна мутация в среднем происходит в гаплотипе за 1/0.12 поколений, то есть примерно раз в 8 условных поколений, или раз в 208 лет. С такими протяженными гаплотипами уже удобно работать, они дают неплохое разрешение при расчетах. Именно так было получено, что 31, 33 и 9 мутаций между парами гаплотипов выше соответствуют временной дистанции 8650, 9400 и 2025 лет, а общий предок жил половину этой временной дистанции назад. При расчетах была сделана поправка на возвратные мутации, которые происходят на больших отрезках времени, превышающих 25 условных поколений, то есть 625 лет.
Теперь понятно, почему гаплотипы выбирают так, чтобы в них таких маркеров было как можно больше (но все-таки чтобы оставаться в рамках практичности), и в ранних работах использовались 6-маркерные гаплотипы, затем 12-маркерные, 17- и 19-маркерные, затем 25- и 37-маркерные, а сейчас работа рутинно ведется с 67- и 111-маркерными гаплотипами (правда, в академических публикациях это обычно от 8 до 17 маркерных гаплотипов). В 111-маркерных гаплотипах одна мутация происходит в среднем за 5 поколений, поэтому у 111-маркерных гаплотипов разрешение лучше других. Но их определять дороже, чем более короткие, поэтому в академических исследованиях, при постоянной нехватке денежных средств, приходится работать с более короткими гаплотипами. Длинные гаплотипы определяют в коммерческих компаниях, обычно персонально, каждый для себя, и обычно передают в общественные базы данных. Сейчас в общественных базах данных — сотни тысяч гаплотипов, и базы прирастают многими гаплотипами ежедневно (Интернет-сайт R1a; Интернет-сайт «A Free Public Service from Family Tree DNA»; Klyosov, Rozhanskii, 2012).
Как мы покажем ниже, эти гаплотипы, а именно числа в них, напрямую связаны с хронологией древних миграций людей, переходами людей на новые места, событиями прошлого — войнами, климатическими катаклизмами, эпидемиями, в общем, со всем тем, что так или иначе влияло на численность популяций человека в определенные времена и на определенных территориях. Например, расчеты могут показать когда древние арии появились на Русской равнине, когда они появились в Индии, Иране, на Ближнем Востоке — потому что эти события приводили к появлению на этих территориях и в те времена общих предков популяций, которые разрастались вплоть до нашего времени, образуя «кусты» гаплотипов со всеми мутациями в них, то есть изменениями чисел в маркерах.
Обратимся к гаплогруппам, тем самым «обложкам паспорта», групповым характеристикам. Относительно умеренное расстояние до общих предков моего и упомянутых киргизских гаплотипов, всего несколько тысяч лет, было вызвано тем, что все мы трое относимся к одной гаплогруппе, то есть к одному роду, под названием R1a. Строго говоря, общий род имеет индекс R, но этот род более 30 тысяч лет назад разошелся на подчиненные рода R1 и R2, а род R1 – примерно 20 тысяч лет назад на рода R1a и R1b. Хотя последние находятся на два уровня ниже рода (гаплогруппы) R, их тоже обычно называют гаплогруппами, хотя правильнее было бы называть субкладами. В целом понятия гаплогруппы и субклада часто взаимозаменимы, и определяются контекстом изложения. И гаплогруппы, и субклады определяются необратимыми мутациями в Y-хромосоме, в отличие от обратимых мутаций в гаплотипах. Эти необратимые мутации, которые называют снип-мутациями (от сокращения SNP, Single Nucleotide Polymorphism), стараются подобрать такие (для идентификации гаплогрупп и субкладов), которые были бы стабильными за все время существования человечества. Поэтому гаплогруппы и субклады стабильны, не меняются во времени.
Всего в мире насчитывают 20 основных гаплогрупп,
которые обозначают буквами латинского алфавита, от А
до Т, хотя систему порой нарушают. В последнее время в классификацию добавили
гаплогруппы A0 и A00, хотя их носителей обнаружили считанные единицы, все они
живут в Африке. Но они настолько отстоят по снип-мутациям от всех остальных
людей на Земле (тестированных на мутации в ДНК), что их пришлось выделить в отдельный род-гаплогруппу. Помимо этого, были идентифицированы промежуточные, сводные гаплогруппы,
такие, как СТ, DE, GHIJK и другие (см. диаграмму
ниже), так что минимальный состав генеалогического дерева мужской половины
человечества включает уже 39 основных гаплогрупп, то есть главных уровней
Y-хромосомного генеалогической структуры. С подгруппами это составляет уже
много сотен.
Если же считать все уровни генеалогического дерева (уровень в этом случае – это субклад), то гаплогруппа R1a, основная гаплогруппа (род) этнических русских, уже насчитывает 43 субклада, гаплогруппа R1b – 177 субкладов, и это при том, что на диаграмме ниже они обе входят в сводную гаплогруппу R, которой примерно 30 тысяч лет со времени образования. Образовалась она при появлении необратимой мутации в виде спонтанного превращения одного нуклеотида в другой в ДНК (Y-хромосоме) патриарха, или его выжившего потомка, в свою очередь потомки которого дожили до настоящего времени в количестве более миллиарда человек. Если точнее, то у него, патриарха гаплогруппы R, по сравнению с ДНК его отца (относящегося к гаплогруппе Р) произошло спонтанное превращение аденина в гуанин, и это произошло в участке Y-хромосомы под номером 15 миллионов 581 тысяч 983. Всего же в Y-хромосоме мужчин насчитывается, как сообщалось выше, примерно 58 миллионов нуклеотидов.
Ствол древнего генеалогического дерева человека
• A00 L1284
• A0-T L1085
• • A0 L991
• • A1 P305, V168
• • • A1a M31
• • • A1b P108,
V221
• • • • A1b1 L419
• • • • BT M91
• • • • • B M60
• • • • • CT M168
• • • • • • DE M145
• • • • • • • D M174
• • • • • • • E M96
• • • • • • CF P143
• • • • • • • C M130
• • • • • • • F M89
• • • • • • • • GHIJK F1329
• • • • • • • • • G M201
• • • • • • • • • HIJK M578
• • • • • • • • • • H L901
• • • • • • • • • • IJK L15
• • • • • • • • • • • IJ M429
• • • • • • • • • • • • I M170
• • • • • • • • • • • • J M304
• • • • • • • • • • • K M9
• • • • • • • • • • • • LT L298
• • • • • • • • • • • • • L M20
• • • • • • • • • • • • • T M184
• • • • • • • • • • • • • NO M214
• • • • • • • • • • • • • • N M231
• • • • • • • • • • • • • • O M175
• • • • • • • • • • • • • • •
• S M230
• • • • • • • • • • • • • • •
M P256
• • • • • • • • • • • • • • P P295,
V231
• • • • • • • • • • • • • • •
Q M242
• • • • • • • • • • • • • • •
R M207
• • • • • • • • • •
• • • • • • R1 M173
• • • • • • • • • • • • • • • • • R1a M420
• • • • • • • • • • • • • • • • • R1b M343
Как получилось так, что русский-славянин и киргизы-тюрки оказались в одном роду, R1a? Причина проста – гаплогруппы намного древнее национальностей, этносов, религий. Примерно 4500 лет назад начались арийские миграции из Европы, видимо, с Балкан, по Русской равнине (Klyosov, 2012a), и между 4000-3500 лет назад арии пришли в Месопотамию (в частности, митаннийские арии, территория нынешней Сирии), в Среднюю Азию и далее на Иранское плато (авестийские арии), и в Индостан (индоарии). Так генеалогически, в одном роду, R1a, роду ариев, оказались и будущие русские, и будущие киргизы, таджики, многие арабы, иранцы, индийцы. Общий их предок в роду R1a жил 5000-4500 лет назад. В настоящее время до 72% состава высших каст в Индии относятся к роду R1a (Sharma et al, 2009).
Рассмотрим киргизов более детально. В сети есть «Киргизский Проект FTDNA» (https://www.familytreedna.com/public/kirgiz/default.aspx?section=yresults).
В нем сейчас (конец декабря 2014 г.) 63 участника, каждый из которых прислал свои результаты тестирования Y-хромосомы, тест проводила компания FTDNA (Family Tree DNA), DNA – это и есть ДНК, в переводе на кириллицу. К сожалению, большинство участников Проекта имеют только 12-маркерные гаплотипы, и очень немногие имеют 67-маркерные. Всего двое определили 111-маркерные гаплотипы. Поэтому рассмотрим дерево 12-маркерных гаплотипов киргизов, для иллюстративной цели этого достаточно. Дерево гаплотипов строится с помощью профессиональной компьютерной программы PHYLIP 3.695 с преобразованием в программе MEGA6. Суть построения дерева в том, что программа группирует индивидуальные гаплотипы по принципу похожести и топологии образования одного гаплотипа из другого. Иначе говоря, получается по сути генеалогическое дерево рассматриваемого набора гаплотипов.
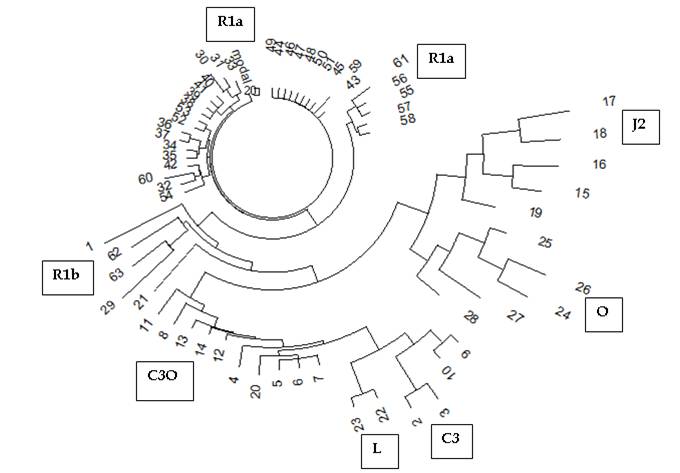
Рис.
1. Дерево 63 гаплотипов в 12-маркерном формате киргизского Проекта FTDNA. Построено
по данным Проекта
https://www.familytreedna.com/public/kirgiz/default.aspx?section=yresults
.
Указаны
гаплогруппы отдельных ветвей.
Мы видим, что среди киргизов, во всяком случае, среди тех, кто сдал ДНК на анализ, наиболее представлены носители гаплогруппы R1a, их 32 человека из 63, то есть 51%. На втором месте – носители гаплогруппы C3, их 14 человек, или 22%. Носителей гаплогруппы О пять человек, то есть примерно 8%. Остальные – единичные представители, которые могут быть случайные в составе киргизов, или как их полушутя называют, «туристы». Это гаплогруппы J2, N1, R1b, I1. В состав данной выборки попали казахи (в Проекте они так и отмечены), и мы видим, что некоторые из них стоят особняком – это гаплотипы 19 (гаплогруппа Q) и 21 (гаплогруппа N), но некоторые относятся к киргизским популяциям (гаплотипы 20 и 50), то есть имеют то же родовое происхождение, что и киргизы.
Это дерево и соответствующие
серии гаплотипов, которые дерево разнесло по ветвям, уже можно количественно
анализировать в отношении хронологии ветвей, то есть времен их происхождения.
Например, основная группа R1a из 27 человек, гаплотипы которых образуют плотную
ветвь в верхней части дерева, имеют следующие 12-маркерные гаплотипы:
13 24 16 10 11 14 12 12 10 13 11 18
13 24 16 11 11 14 12 12 10 13 11 18
13 24 16 11 11 14 12 12 10 14 11 17
13 24 16 11 11 14 12 12 10 14 11 18
13 25 16 10 11 14 12 12 10 14 11 18
13 25 16 10 11 14 12 12 10 14 11 18
13 25 16 10 11 14 12 12 10 14 11 19
13 25 16 10 11 14 12 12 10 14 11 19
13 25 16 11 11 14 12 12 10 13 11 18
13 25 16 11 11 14 12 12 10 13 11 18
13 25 16 11 11 14 12 12 10 13 11 18
13 25 16 11 11 14 12 12 10 13 11 18
13 25 16 11 11 14 12 12 10 14 11 17
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 18
13 25 16 11 11 14 12 12 10 14 11 19
13 25 16 11 11 14 12 12 10 14 11 19
13 25 16 11 11 14 12 12 10 15 11 17
13 25 17 11 11 14 12 12 10 14 11 18
13 26 16 11 11 11 12 12 10 14 11 17
Девять из них идентичны друг другу, как дерево гаплотипов и показывает (девять одинаковых коротких палочек на вершине дерева) – это базовые, или предковые гаплотипы. Они не успели мутировать за время, прошедшее от их общего предка. На все 27 гаплотипов приходится 27 мутаций (отмечены), что дает 27/27/0.02 = 50 à 53 условных поколения, то есть 1325±290 лет до общего предка. Это – примерно 7-й век нашей эры, плюс-минус три века. Погрешность определяется по обычным правилам математической статистики. Значительная погрешность, несколько более 20%, вызвана тем, что в серии всего немного гаплотипов, и все короткие, да и мутаций мало. Были бы несколько сотен гаплотипов, да 67-маркерных, то и надежность расчетов была бы намного выше. Но и здесь представление о временах жизни общего предка данной серии гаплотипов неплохое, это скорее всего скифы-сарматы и их не очень удаленные потомки. В расчетах выше 0.02 – это константа скорости мутации в 12-маркерных гаплотипах (выраженная в числе мутаций на условное поколение в 25 лет), стрелка – поправка на возвратные мутации, которая или рассчитывается по определенной формуле, или получается по опубликованным таблицам (Клёсов, 2008b). Этот способ расчета в ДНК-генеалогии называется линейным.
Проверить расчеты можно с
помощью логарифмического метода (Klyosov, 2009a), даже не считая мутации.
Поскольку в серии из 27 гаплотипов 9 базовых, то получаем [ln(27/9)]/0.02 = 55 à 58 условных поколений, то
есть 1450±500 лет до общего предка. Как видно, это в пределах погрешности
расчетов совпадает с величиной, полученной линейным способом. Для
концептуальных выводов это вполне приемлемо.
Но мы видим, что на дереве
есть еще одна небольшая ветвь из пяти гаплотипов гаплогруппы R1a, справа
наверху, причем ветвь, не выходящая из предыдущей, а
независимая. Ясно, что общий предок у обеих ветвей был, поскольку гаплогруппа
одна и та же, но предок второй ветви определенно более древний (потому что
ветвь более раскидистая). Взглянем на эти гаплотипы:
13 25 16 11 11 14 12 12 11 14 11 17
13 25 16 11 11 14 12 12 11 14 11 17
13 25 16 11 11 14 12 12 11 14 11 18
13 25 16 11 11 14 12 12 11 14 11 18
13 26 16 11 11 14 12 12 11 14 11 17
У них два базовых гаплотипа из пяти, и всего три мутации от базового гаплотипа на всех. Получаем, линейный метод: 3/5/0.02 = 30 à 31 условных поколений, то есть 775±450 лет до общего предка. Логарифмический метод для таких малых серий применять практически бессмысленно, но попытаемся: [ln(5/2)]/0.02 = 46 à 48 условных поколений, то есть 1200±860 лет до общего предка. Результаты совпали в пределах погрешности, но пользы от этого мало. Потому что там, где всего два базовых гаплотипа, там легко могут оказаться один или три, мутации-то неупорядоченные, статистические. При трех базовых гаплотипах датировка оказалась бы 675 лет назад, при одном базовом – 2175 лет назад, тем более что «один базовый гаплотип» - понятие само по себе бессмысленное. Поэтому в таких ситуациях намного предпочтительнее линейный метод расчета.
Еще важное наблюдение – базовые гаплотипы у двух рассмотренных ветвей различаются на две мутации:
13 25 16 11 11 14 12 12 10
14 11 18
13 25 16 11 11 14 12 12 11 14 11
17
На самом деле там не две мутации разницы, а всего 0.91 мутации, поскольку средние величины дробные. Это разводит общих предков двух данных ветвей на 0.91/0.02 = 46 à 48 условных поколений, то есть на 1200 лет, и их общий предок жил примерно (1200+1325+775)/2 = 1650 лет назад, примерно в 4-м веке нашей эры. Это, наиболее вероятно, все те же скифы или сарматы. Но здесь ДНК-генеалогия должна согласовать свои предположения с исторической наукой, и либо ее дополнить, либо, напротив, откорректировать свои предположения.
Как мы видим, мутации в
гаплотипах потомков расходятся от предкового гаплотипа
как круги по воде, число мутаций легко рассчитывается, и они подчиняются
довольно простым количественным закономерностям. Для кругов на воде,
расходящихся от места, куда был брошен камень, легко рассчитать, когда был
брошен камень, если знать скорость распространения волны и место нахождения круговой
волны в данный момент времени. Чем больше прошло времени – тем дальше круги
ушли, тем больше они разошлись. Так и в гаплотипах - чем больше время,
прошедшее от общего предка, тем больше мутаций накопилось в гаплотипах его
потомков. Число этих мутаций связано с временем,
прошедшим от общего предка, с числом гаплотипов в серии, и с константой
скорости мутации в гаплотипах, и выражается простой формулой: n/N = kt, где n –
число мутаций в серии из N гаплотипов, k – константа скорости мутации (в числе
мутаций на гаплотип за условное поколение, равное 25 лет), t – число условных
поколений, с табличной поправкой на возвратные мутации (Klyosov, 2009а). На
сотнях и тысячах примеров показано, что эта формула работает при любом числе
гаплотипов и мутаций в них, и при любом времени, прошедшем от общего предка
рассматриваемых гаплотипов. Однако при очень больших временах, более 10-20
тысяч лет, и особенно более 100 тысяч лет, нужно использовать гаплотипы с
«медленными» маркерами, то есть с малыми константами скоростей мутаций, и тем
самым снижать число мутаций и число возвратных
мутаций. По аналогии, вряд ли целесообразно изучать скорости радиоактивного
распада элементов со временами полураспада в тысячелетия, используя секундомер.
Или пытаться изучать круги на воде за километры от места, куда был брошен
камень, для этого нужно значительно более мощное воздействие. Как всегда, нужен
конкретный анализ в конкретной ситуации, единых подходов на все случае жизни не
бывает. Варианты конкретного анализа в конкретных ситуациях и рассматривает
ДНК-генеалогия. Некоторые ситуации и расчеты мы расмотрим ниже.
Как мы видим, мутации в
ДНК-генеалогии – это не только единичные необратимые мутации-снипы, описанные
выше, которые определяют гаплогруппы и субклады, но и обратимые мутации,
меняющие числа повторов, или аллели, в гаплотипах. В русскоязычной литературе
их называют просто «мутации», с пониманием, что это не те мутации (в генах),
которые обычно возникают под действием радиации. Переход числа повторов в
маркере Y-хромосомы от 25 к 24 или 26 (или наоборот) имеет совершенно другую природу,
чем «поломка» гена. Такой переход является следствием спонтанной ошибки
ДНК-копирующей «биологической машины», это процесс первого порядка с точки
зрения физико-химической кинетики, он не зависит от внешних воздействий. У предка киргизов гаплогруппы R1a второе
число указывает на 25 повторов четверки TCTG/TCTA, гаплогруппы R1b - 19
повторов в том же маркере – это древняя сибирская (по происхождению)
гаплогруппа/субклад R1b-M73. Гаплогруппа C3 у киргизов по происхождению монголоидная, прибыла, видимо, из монгольских степей. С
востока – и гаплогруппа О, основная гаплогруппа в
Китае. Оттуда же – гаплогруппа N1, которая со стороны
Китая и затем Алтайского региона прошла на север против часовой стрелки, и
через северный Урал достигла Прибалтики, став финно-уграми (один вид гаплотипов
и одни субклады, к тому же язык финно-угорский) и южными балтами (другой вид
гаплотипов и другие субклады, язык в основном индо-европейский). Но малой
частью носители гаплогруппы N1 попали в Среднюю Азию, и к киргизам. Гаплогруппа
Q – опять сибирская, гаплогруппа J2 – напротив,
прибыла со стороны Средиземноморья, но в Киргизию – из Ирана или Индии, где их
много.
После этого короткого
экскурса в практическую ДНК-генеалогию рассмотрим предмет более основательно.
Введем некоторые важные понятия и определения. Ниже они будут пояснены более
детально, но важно их сразу обозначить. Во-первых, понятие «ДНК-генеалогия».
Это – не генетика, в чем часто путаются даже професионалы в науке. Генетика
человека – это наука, изучающая наследственность и изменчивость признаков,
определяющих врожденные особенности человека, и передаваемых, как правило,
через гены. Но ДНК только на 2% состоит из генов, а в Y-хромосоме генов вообще
ничтожная доля. Но дело даже не столько в этом. Направления и области науки
определяются не объектами исследования, а методологией исследования. Именно
методология исследования отличает, например, химию от физики, хотя объекты
часто одинаковы. Методология генетики совершенно другая, чем методология
ДНК-генеалогии. У них разные задачи исследований. ДНК-генеалогия
по сути историческая наука, она оперирует хронологией, датировками древних событий,
и для этого ДНК-генеалогия использует свой расчетный аппарат, которого нет в
методологии генетики. ДНК-генеалогия использует и развивает методы
физико-химической кинетики в применении к ДНК, переводит картину мутаций в
хронологические, исторические показатели, как указывалось выше. Это вообще не
входит в методологию генетики, у нее другой экспериментальный и аппаратурный
базис.
Ошибка – полагать, что
направление науки определяется объектом исследования. Изучать ДНК – это далеко
не обязательно генетика. Например, химик, растворяя ДНК в кислоте и изучая,
скажем, вязкость получаемого раствора, вовсе не занимается генетикой. Альберт
Сент-Дьорди как-то сказал – «Дайте химику динамомашину, и он тут же растворит
ее в соляной кислоте». Это вовсе не означает, что химик при этом будет
заниматься электродинамикой.
Некоторые, не слишком
знакомые с предметом, полагают, что ДНК-генеалогия – это популяционная
генетика. Это – совершенно не так. Популяционная генетика – это часть генетики,
о чем говорит ее название, но со своими особенностями. Главная задача
популяционной генетики – это выявление связи между генотипом и фенотипом, то, к
чему ДНК-генеалогия не имеет отношения. Популяционная генетика зачастую тоже
рассматривает гаплогруппы и гаплотипы Y-хромосомы, как и ДНК-генеалогия, но на
этом сходство заканчивается. Аппарат популяционной генетики, применяемый для
интерпретации получаемых данных, например, «метод главных компонент»,
характеризуется со стороны ДНК-генеалогии как совершенно примитивный и ведущий
к заблуждениям, и не используется в ДНК-генеалогии. Он не отвечает задачам
ДНК-генеалогии. Это же относится и к искаженным и примитивным методам обработки
картин мутаций в ДНК в рамках популяционной генетики. Если
популяционный генетик увидит (или получит) набор из сотни гаплотипов, скажем,
37-маркерных, в котором, например, четыре гаплотипа одинаковых, и еще семь
будут совпадать друг с другом случайными парами, он сообщит, что в наборе
имеется «89 уникальных гаплотипов», и это и будет результатом его исследования,
который пойдет в статью в научный журнал. Ни он, ни рецензенты не хотят
признать, это эта «информация» не имеет ни малейшей ценности, и фактически
никому в таком виде не нужна. Но таковы принципы и правила популяционной
генетики. «Молекулярной историей» это назвать никак нельзя. А
специалист в ДНК-генеалогии сразу скажет, что общий предок этих ста гаплотипов
жил 925±105 лет назад, потому что натуральный логарифм отношения 100/4,
поделенный на константу скорости мутации для 37-маркерных гаплотипов (0.09
мутаций на гаплотип на условное поколение в 25 лет) равен 36 à
37 условных поколений, то есть общий предок этой сотни гаплотипов жил примерно
925 лет назад. Сказать это с большей определенностью можно тогда, когда
подсчитано число мутаций во всех ста гаплотипах, и оно будет равно 324, и тогда
324/100/0.09 = 36 à 37 условных поколений, то
есть ровно тот же промежуток времени до общего предка. Погрешность расчетов
определяется по известным правилам (Klyosov, 2009a). Совпадение времен до общего предка для
«логарифмического» и «линейного» метода (в первом мутации не считаются, во
втором считаются) свидетельствует, что закономерности образования мутаций в гаплотипах
следуют кинетике первого порядка, что в свою очередь означает, что все сто
гаплотипов действительно произошли от одного прямого общего предка.
Популяционная генетика такими подходами не пользуется, они ей чужды по ряду
причин, среди них – приверженность устаревшим, традиционным подходам,
отсутствие соответствующего образования, клановость, отсутствие хорошей научной
школы. Короче, ДНК-генеалогия является другой дисциплиной, нежели популяционная
генетика.
ДНК-генеалогия базируется на
четких правилах, которых можно насчитать четырнадцать.
Первое — как уже было отмечено, к
генетике ДНК-генеалогия прямого отношения не имеет. ДНК (дезоксирибонуклеиновая
кислота) — это не только гены. В ДНК есть протяжённые последовательности повторяющихся
нуклеотидов, в которых, повторяем, генов нет (поэтому поначалу эти
последовательности назвали «никчемными»), и эти последовательности детально
копируются при передаче наследственной информации от отца к сыну. По мере
исследований, оказалось, что эти последовательности хранят массу информации и
не только «генеалогического» характера. Эти негенные участки ДНК в мужской
Y-хромосоме, одной из 23 хромосом, которые доставляются сперматозоидом в
организм будущей матери, копируются от отца к сыну, поколение за поколением.
Поскольку у матери такой хромосомы нет, то сын наследует её только от отца.
Хромосомы сплетаются с материнской ДНК и образуют новую комбинированную
молекулу ДНК, в которой хромосомы отца и матери перетасовываются.
Перетасовываются все, кроме мужской Y-хромосомы.
Так и получается, что отец
передаёт сыну эту Y-хромосому интактной, строго скопированной со своей. Свою Y-хромосому он получил от своего отца. Тот —
от своего. И так далее, на тысячи, десятки тысяч и сотни тысяч лет вглубь,
назад, к далёким предкам современных людей и к предкам тех далёких предков.
Если бы
Y-хромосома так и передавалась из поколения в поколение действительно
неизменной, толку для генеалогии от неё было бы мало. Но неизменного ничего в
мире нет, особенно когда речь о копировании. Копирования без ошибок не бывает.
В том числе и копирования Y-хромосомы.
Отсюда второе положение — время от времени при
копировании Y-хромосомы в копии проскакивают ошибки. Фермент
под названием ДНК-полимераза (на самом деле вместе с целым набором молекулярных
инструментов) или просто ошибается и делает некоторые участки ДНК короче или
длиннее, удлиняя или сокращая повторы нуклеотидов, или «ремонтирует»
повреждённые участки и в ходе «ремонта» удаляет повреждение (эта операция
называется «делеция») или «вшивает» новый нуклеотид (называется «вставка», или
«инсерт»). То, что удалось отремонтировать, в мутации не попадает, как
будто мутации и не было. Что не удалось — передается сыну при рождении. Если
родилась дочь, то Y-хромосома не передается. Если только дочери, или детей нет
— Y-хромосома терминируется, вместе с мужской наследственной линией.
Мутации, которые интересуют
ДНК-генеалогию, бывают двух типов — или изменение числа повторов, тандемов
(STR), что описано выше, или «точечные», одно- или несколько-нуклеотидные
(SNP).
Оказалось, что многие
повторы нуклеотидов находятся у разных людей в одних и тех же участках
Y-хромосомы. Эти участки уже специалистами пронумерованы, классифицированы,
сведены в списки. Как было описано выше, они получили название «маркеры». Набор
маркеров, точнее, повторов в них, называется «гаплотип». Уже известных маркеров — более тысячи. Они, как правило,
одни и те же у всех людей на Земле и отличаются друг от друга только числом повторов,
что вызвано теми самыми ошибками ферментов при копировании из поколения в
поколение. Внимательное рассмотрение маркеров и гаплотипов позволило сделать
вывод, что все люди на Земле произошли (точнее, потомки выжили) от одного
общего предка. Но здесь понятие «одного предка» можно рассматривать на разных
уровнях сложности. Например, древнейшая африканская линия (гаплогруппа А00)
имеет общего предка с гаплогруппой R1a примерно 210 тыс. л.н., то есть оттуда,
из такой глубины тысячелетий расходятся гаплотипы современных носителей
гаплогрупп A00 и R1a:
13 11 12 10 11 16 10 8 14 14 8 8 8 9 12 11 12 8 12 12 11 11 (А00)
12 12 11 11 11 11 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a)
Здесь приведены 22 очень «медленных» маркера Y-хромосомы, каждый из которых мутирует раз во многие тысячелетия. Можно подсчитать, что эти гаплотипы разошлись на 26 мутаций, и расчеты, основанные на скорости мутации каждого маркера, показывают, что эти 26 мутаций соответствуют времени жизни их общего предка 210 тыс. л.н.. Но даже беглый взгляд на эти гаплотипы показывает, что ДНК древнейших африканцев и этнических русских, носителей гаплогруппы R1a, не так уж намного различаются. В рамках скоростей медленных 22-маркерных гаплотипов, они разошлись всего 160 тысяч лет назад.
Хорошо, а если вместо типичного гаплотипа R1a подставить гаплотип R1b, преобладающей
гаплогруппы Западной Европы? Мутации здесь настолько медленные, что в таком
формате представления гаплотипа он будет практически одинаков по всей Западной
Европе:
11 12 13 11 11 12 11 9 15 16 8 10 8 12 10 12 12 8 12 11 11 12 (R1b)
Мы видим, что при таком разрешении большая часть аллелей (это — числа в гаплотипах) у R1a и R1b одинаковы (совпадают 16 аллелей из 22). Между А00 и R1b — 24 мутации, что снижает возраст общего предка на семь тысяч лет, то есть на 3%. Это — в пределах погрешности расчетов. Только не стоит строить пропорции, там счет нелинейный.
Можно ли считать, что общий
предок человечества жил 200-210 тыс. л.н.? Вряд ли, поскольку гаплогруппа А00
была обнаружена всего два года назад, и до этого считали, что общий предок
человечества жил 140 тыс. л.н. Если завтра исследователи найдут новую гаплогруппу, то срок начала человечества опять
отодвинется. И он несомненно будет отодвигаться
дальше, по мере увеличения наших знаний. Уже появилась еще одна реперная точка
— это общий предок шимпанзе и человека. У современного шимпанзе из полной
хромосомной последовательности удалось определить 16 маркеров Y-хромосомы
(Klyosov et al, 2012). Они показаны рядом со значениями аллелей в тех же
маркерах Y-хромосомы человека:
8 15 10 4 5 9 10 5 10 4 4 7 4 4 8 9 (шимпанзе)
11 12 11 11 10 8 10 8 12 10 12 12 12 11 11 11 (человек)
Это оказались еще более медленные маркеры, и 64 мутации (!) между ними помещают общего предка шимпанзе и человека на глубину времен в 5.5 миллионов л.н.. В целом это же дают и антропологические оценки. Что важно — никаких «инопланетян» в создании современного человека не было, вместо этого имеется непрерывный эволюционный путь от общего предка шимпанзе и человека в ходе миллионов лет к нам, современным людям.
Итак, положение третье:
все люди — родственники, они все происходят в целом от одного предка, хотя его
идентификация зависит от определений, кого им считать. Этот предок оказался
древнее, чем предполагалось раньше, но он был. Еще недавно
считалось, что он жил примерно 70 тыс. л.н., потом 120 тысяч, потом 140 тысяч,
сейчас он уже уходит глубже 200 тыс. л.н., постепенно приближаясь к общему
предку с неандертальцем 300-500 тыс. л.н., и, возможно, уйдет еще далее к
приматам, миллионы л.н.. Как мы видим, гаплотипы хоть и отдаленно, но в
принципе похожи на те, что есть у всех нас. У шимпанзе и у нас — одни и
те же маркеры. Следовательно, общий предок был один и тот же, только очень
древний.
Положение четвёртое
— гаплотипы изображают в виде числа тандемов, или повторов, по каждому маркеру,
выбранному из десятков и сотен. В англоязычной литературеих
называют STR, или Short Tandem Repeats. Самый простой и короткий гаплотип из
тех, которые рассматривает ДНК-генеалогия, состоит из пяти или шести маркеров.
Например, у носителей гаплогруппы R1a, основной в Киргизии, как сообщалось
выше, базовый (или предковый) 6-маркерный гаплотип, в
той записи, которую обычно приводят в научных публикациях, особенно в
устаревших, такой:
16-12-25-11-11-13
у носителей гаплогруппы R1a-Z280, типичной
для современных этнических русских, 6-маркерный гаплотип точно такой же:
16-12-25-11-11-13
у носителей R1a-L342.2, наиболее полно соответствующих
степным ариям III тыс. до н.э., опять точно такой же:
16-12-25-11-11-13
у скифов из раскопок в Минусинской котловине, с
датировками 3800-3400 л.н., опять точно такой же:
16-12-25-11-11-13
в высших кастах Индии (R1a-L342.2-L657), опять точно
такой же:
16-12-25-11-11-13
у западных славян гаплогруппы R1a-M458, которая образовалась на территории Белоруссии-Украины
примерно 4200 л.н., уже после ухода ариев с Русской ранины в Месопотамию
(митаннийские арии), на Иранское плато (авестийские арии), на Южный Урал и затем
в Индостан (индоарии),
17-12-25-10-11-13.
Потомки древних носителей гаплогруппы R1a-M458 сейчас живут в основном в Белоруссии, Польше, на Украине, продвинулись в Центральную Европу; возможно, это они были первыми кельтами в Центральной Европе (в частности, в Австрии, образуя ядро гальштаттской археологической культуры). Как мы видим, 6-маркерный гаплотип уже другой, и легко отличается.
У киргизов, носителей гаплогруппы С3, 6-маркерный гаплотип следующий:
15-14-25-10-11-13
Мы видим, что даже короткие, 6-маркерные гаплотипы
являются вполне информативными, и позволяют сразу распознать представителей
арийских народов и их потомков — скифов, восточных славян, состав высших каст в
Индии (гаплогруппы R1a, которая там доминирует). Более протяженные гаплотипы,
вплоть до 67- и 111-маркерных, позволяют уточнить картину, выявить более
недавние ДНК-генеалогические линии, определить их
датировку. Но суть от этого принципиально не меняется.
Положение пятое
— по числу мутаций в гаплотипах можно определять возраст гаплотипа, то есть
время, прошедшее от общего предка этого гаплотипа до его сегодняшних потомков. Поскольку сыновья сохраняют гаплотип отца, переданный по
наследству, и мутации в этом гаплотипе проскакивают в среднем только раз
примерно в 3100 лет (6-маркерный гаплотип) или раз в 1300 лет (12-маркерный
гаплотип), или раз в 125 лет (111-маркерный гаплотип), то даже через 5000 лет у
потомков сохранится 23% исходного 6-маркерного гаплотипа, без изменений.
То есть в списке из 100 гаплотипов потомков — 23 гаплотипа будут такими же,
какой был у предка 5000 л.н.. Это если рассматривать
6-маркерные гаплотипы. При рассмотрении 12-маркерных гаплотипов те же 23%
гаплотипов предка сохранятся через 88 поколений, или 2200 лет.
Таким образом, определить
гаплотип предка можно и через тысячелетия. И по его виду можно узнать, из каких
краёв предок пришёл, сравнив вид гаплотипа с гаплотипами по территориям, и с
доступными ископаемыми гаплотипами.
Положение шестое
— гаплотипы не указывают на этносы, это совершенно разные понятия. Гаплотипы
указывают на древние рода, племена, которые намного старше этносов. С тех
древних времен гаплотипы давно разошлись по разным территориям, на которых
потом, через тысячелетия сформировались этносы.
Положение седьмое
- доля в процентах гаплогруппы в популяции сама по себе означает не очень
много, это просто структура современной популяции, то, чем занимаются
популяционные генетики. Часто бывает, что доля большая, а общий предок недавний,
просто потомки общего предка быстро размножились, условия были благоприятными.
Большая доля гаплогруппы вовсе не указывает на ее «прародину». Например, доля
гаплогруппы R1b в Ирландии достигает 90%, а гаплогруппа R1b зародилась в Южной
Сибири. Доля гаплогруппы R1a в высших кастах Индии достигает 72%, а сама
гаплогруппа пришла в Индию с ариями примерно 3500 лет назад.
Поскольку у каждого племени
и в каждом этносе, в каждом регионе можно определять возраст племени и вообще
популяции в целом, то можно определять, когда и в каком направлении шли древние
миграции.
Положение восьмое
— все расчеты производятся с погрешностями. Точные цифры получены быть не могут
в принципе. Дело в том, что мутации неупорядоченные, поэтому мы имеем дело со
статистикой (Klyosov, 2009a). Чем больше выборка, тем точнее расчеты. Чем
длиннее гаплотипы — тем точнее расчеты. Погрешность зависит от числа гаплотипов
в выборке, от протяженности гаплотипов, от того, насколько точно определена и
выверена, откалибрована константа скорости мутации, от того, насколько древний
общий предок. Особенно неточно сравнивать мутации в парах гаплотипов людей. На
двух гаплотипах мутация-другая могла добавиться буквально в предыдущем или
нынешнем поколении. Это может сразу привести к прибавлению-отнятию тысячи лет
«в одном поколении». А в большой выборке разница относительно нивелируется
статистикой. Есть выборки в тысячи гаплотипов — там расчеты, конечно, точнее.
В ДНК-генеалогии часто важна
не абсолютная точность, а концептуальный вывод. Например,
если некто утверждает, что носители гаплогруппы R1b (которые сейчас составляют
примерно 60% мужского населения Западной и Центральной Европы), жили там, в
Европе, еще 30 тыс. л.н. (а такими утверждениями, без расчетов, была заполнена
академическая литература по популяционной генетике до недавнего времени), а
расчеты показали 4800 л.н., то здесь дело не в точности, а в принципиальном
выводе. То же самое по ДНК-генеалогии Кавказа — если
данные показывают, что Кавказ заселялся носителями гаплогруппы J2 из Месопотамии
примерно 7-6 тыс. л.н., причем заселялся разными племенами и по разным
территориям, и уже известно каким именно, и в какое время, то это дает важные
данные историкам и археологам, которые продолжают горячо спорить по данным
вопросам. То же самое по ДНК-генеалогии Прибалтики —
данные показывают, что Прибалтика со стороны Финляндии и со стороны Южной
Балтики заселялась всего 2000-2500 л.н. (со стороны Южной Балтики примерно на
500 лет раньше), но сами мигранты, которые на исходе миграции разделились на
две большие ветви — северная и южная — имеют общего предка примерно 3575 л.н.,
который, видимо, жил на Урале (Клёсов, А.А., Пензев
К.А. Арийские народы на просторах Евразии, 2014).
К этому
ведёт положение девятое — гаплотипы в немалой степени (но не всегда) связаны с
определёнными территориями.
Но как такое может быть? А
вот как. В древности большинство людей передвигались племенами, родами. Род, по
определению, это группа людей, связанных родством. То есть
гаплотипы у них одинаковые или близкие. Помните — одна мутация в среднем
происходит за тысячелетия? Проходили тысячелетия, численность родов порой
сокращалась до минимума («бутылочное горлышко» популяции), и если выживший имел
некоторую мутацию в гаплотипе, то его потомки уже «стартовали» с этой новой
мутацией, копируя ее поколениями в своих ДНК, а мужчины — в своей Y-хромосоме.
В популяционной генетике это называется «эффект основателя», что, в общем,
особой дополнительной смысловой нагрузки не несет. Некоторые люди покидали род
по своей или чужой воле — плен, бегство, путешествия, военные походы, и
выжившие начинали новый род на новом месте. В итоге карта мира с точки зрения
ДНК-генеалогии получилась пятнистой, и каждое пятно порой имеет свой
превалирующий гаплотип — гаплотип рода. Часто он и есть гаплотип предка,
начавшего род на данной территории.
Но есть ещё один тип мутации
— точечные мутации, снипы (калька с английского SNP, что в примерном переводе и
означает «единичная нуклеотидная мутация», или «единичная нуклеотидная
вариация»). Они — практически вечные. Раз появившись, они уже не исчезают.
Теоретически, в том же нуклеотиде может произойти другая мутация, изменив
первую. Но нуклеотидов — миллионы, и вероятность такого события крайне мала.
Всего в хромосомах имеются многие миллионы снипов, из которых в ДНК-генеалогии
применяются более тысячи, и каждый снип соответствует гаплогруппе, то есть
роду, или субкладу, то есть племени, хотя эти дефиниции применяют весьма
вольно, как пояснено выше. Двадцати наиболее крупным гаплогруппам, каждая из
которых охватывает сотни миллионов людей, присвоили буквенные обозначения от А до Т, примерно в хронологическом порядке появления
соответствующего рода на планете. Или, по крайней мере, в том порядке, как
учёные полагают эти рода появились. Хотя ревизий на этот счёт предстоит ещё
много. Как уже упоминалось выше, недавно прошла очередная — в список добавили
две гаплогруппы, А00 и А0, которые недавно обнаружили
в Африке, и которые стоят совершенно особняком даже к другим африканцам, не
говоря о европейцах или азиатах.
Итак, положение десятое
— людей можно классифицировать по древним родам не только (и не столько) по
виду гаплотипов, но и по наличию определённых снипов. Например, носители
гаплогрупп А и В сейчас в основном живут в Африке (но где
эти гаплогруппы появились, пока непонятно, и скорее всего не в Африке);
гаплогруппа С встречается особенно часто среди монголоидов и жителей Австралии
и Океании, хотя жители Австралии и Океании уже далеко не монголоиды, подгруппы
(субклады) далеко отклонились друг от друга в ходе развития или эволюции); гаплогруппу G находят в древней Европе (в основном ископаемые
гаплотипы), на Кавказе, в Передней Азии; гаплогруппа J — исходно Ближний Восток
и оттуда перешла Кавказ не позднее 6-7 тыс. л.н.; представителей гаплогруппы J1
довольно много на Кавказе, а также среди арабов и евреев на Ближнем Востоке,
гаплогруппы J2 много на Кавказе, в Месопотамии и среди жителей Средиземноморья;
гаплогруппа L появилась, похоже, в Передней Азии, и разошлась в разные стороны
— ее подгруппа (субклад) L1a — в Индии, L1b — в Грузии, L1c — на восточном
Кавказе. Гаплогруппу N имеют многие жители Китая, Сибири,
севера России, Прибалтики и части Скандинавии, куда и прибыли носители
гаплогруппы N с востока — Урала и до того Южной Сибири тысячелетия назад).
Гаплогруппа I — возможно,
имеет историю на Русской равнине продолжительностью не менее 40-45 тысяч лет, и
после почти полного вымирания или истребления примерно 4500 л.н. сейчас, почти
исключительно, находится в Европе, где возродилась относительно недавно,
3600-2300 л.н. (для I1 и I2, соответственно); гаплогруппы R1a и R1b появились в
Южной Сибири (примерно 20 тысяч лет назад) после долгой миграции их предков,
носителей гаплогруппы К, из которой образовалась гаплогруппа Р
и затем Q и R, возможно, из Европы или с Русской равнины на восток, далеко за
Урал. Оттуда гаплогруппы R1a и R1b пришли в Европу, причем разными
миграционными путями. R1a стала основной гаплогруппой Восточной Европы, R1b —
западной и центральной Европы.
Данное положение сводится к
тому, что у каждого мужчины есть снип из определённого набора, по которому
можно отнести носителя к определённому древнему роду. Времена появления снипов,
обычно применяемых в ДНК-генеалогии, относятся к диапазону от 25-40 тысяч лет
для «старых» снипов до 10-15 тысяч лет для «молодых», но сейчас уже выявляют
снипы, образовавшиеся всего 1000-1500 л.н.. Например, четверть всего
ирландского мужского населения имеют снип (R1b-M222), который образовался около
2000 л.н., в начале нашей эры В континентальной Европе
его практически нет.
Положение одиннадцатое
— гаплогруппы не просто соответствуют определённым родам, но образуют
определённую последовательность, лестницу гаплогрупп, показывающих их иерархию
— последовательный, ступенчатый переход от точки расхождения африканских
популяций и неафриканских (примерно 160 тыс. л.н.), которые друг от друга не
происходят, до самой недавней гаплогруппы R, образовавшейся примерно 30-40 тыс.
л.н.. Эта лестница называется филогенетическим деревом гаплогрупп и их снипов. Все гаплогруппы и субклады
на дереве должны включать снипы «вышестоящих» гаплогрупп и субкладов. То есть
принцип «лесенки» должен выполняться. Преемственность узловых родов
человечества должна соблюдаться.
Всё это делает
филогенетическое дерево гаплогрупп достаточно прочной и обоснованной
структурой. У него есть, впрочем, слабое место — его филогения не показывает,
на каком континенте зародилось человечество, откуда пошли гаплогруппы, начиная
с первых, на общем стволе, идущим от нашего общего предка с современным
шимпанзе. Говоря языком филогении, дерево гаплогрупп не «укоренено». Укоренение
дерева — результат интерпретаций наблюдений и доступных экспериментальных
данных.
Положение двенадцатое
— в ДНК-генеалогии обычно оперируют поколениями. Поколение в контексте
ДНК-генеалогии — это событие, которое происходит четыре раза в столетие.
Численно и по времени оно близко к продолжительности поколения в житейском
смысле этого слова, но не обязательно равно ему. Хотя бы потому, что продолжительность
«бытового» поколения не может быть точно или хотя бы в среднем определена, она
«плавает» в реальной жизни в зависимости от многих факторов, включая культурные,
религиозные и бытовые традиции, примерно и в среднем от 18 до 36 лет, хотя границы
этого не определены.
В древности этот диапазон
был, видимо, заметно смещён к первой величине. Поэтому использовать столь
«плавающую» величину для расчётов в широких временных диапазонах и для разных народов
не представляется возможным или разумным. Исходя из этого положения, скорости
мутаций откалиброваны под условно взятое поколение продолжительностью 25 лет (Klyosov,
2009a). Если кому-то больше нравится 30 лет на поколение или любое другое
количество лет, скорости можно перекалибровать, и в итоге окажутся ровно те же
величины в годах. Так что сколько лет приходится на
поколение — в данном случае не имеет значения, потому что при расчётах меньшему
числу лет на поколение будет просто соответствовать пропорционально большее
число поколений, и итоговая величина в годах не изменится.
Положение тринадцатое
— только те мутации в гаплотипах имеет смысл рассчитывать, экстраполируя ко
времени общего предка, которые подчиняются определённым количественным закономерностям.
Другими словами, ДНК-генеалогия оперирует тремя
экспериментальными факторами: 1) наличием снипов, относящих человека к определённому
роду; 2) наличием мутаций, позволяющих оценивать время, прошедшее от общего
предка совокупности гаплотипов и — при больших выборках — от начала самого
рода, от самого далёкого из предков ныне живущих потомков данного рода (то есть
здесь считаются сами мутации); 3) закономерностями переходов гаплотипов
в их мутированные формы, без численного учёта самих мутаций (то есть здесь
мутации не считаются, считаются немутированные гаплотипы).
Это позволяет оценить,
насколько достоверны расчёты предка по мутациям, и даёт ещё один, независимый
способ расчетов. Методы, в которых считаются мутации, называются линейным,
квадратичным и пермутационным (Klyosov, 2009a), из которых наиболее прост
линейный метод. Метод, в котором считаются немутированные гаплотипы, называется
логарифмическим. В принципе, он использует формулы химической кинетики первого
порядка. В линейном методе общее число мутаций в серии гаплотипов делится на число
гаплотипов и на константу скорости мутации для данных гаплотипов. В логарифмическом
методе берется логарифм отношения общего числа гаплотипов к числу немутированных
гаплотипов, и делится на константу скорости мутации.
Положение четырнадцатое
— в большинстве случаев результаты расчетов почти не зависят от размера выборки
(при числе гаплотипов больше двух-трех десятков), то есть они устойчивы к
статистическим вариациям. Размер выборки увеличивает точность, и то только до
определенных пределов. Это относится к довольно большим популяциям, которые
перемешались за тысячелетия, но именно с такими обычно
и работают.
Приведем пример. В таблице
ниже приведены расчеты времени до общего предка гаплогруппы R1a на постсоветском
пространстве. Это, в основном, — Россия, Украина, Белоруссия,
Литва, Латвия, Эстония и Казахстан. Видно, что при увеличении выборки от
26 гаплотипов до более шестисот результаты практически одинаковы в пределах
погрешности расчетов.
Таблица 1. История
определений времени до общего предка гаплогруппы R1a1 на «пост-советском
пространстве» по разным сериям гаплотипов (Клёсов, 2011).
|
Дата. |
Число гаплотипов. |
Общее число мутаций. |
Время до общего предка, годы. |
Ссылка. |
|
Июнь 2008 |
26 |
178 |
4400±550 |
Вестник Академии ДНК-генеалогии, т.1 400-477 (2008) |
|
Ноябрь 2008 |
44 |
326 |
4825±550 |
Вестник Академии ДНК-генеалогии, т.1 947-957 (2008) |
|
Январь 2009 |
58 |
423 |
4725±520 |
J. Genetic Genealogy, 5, 186-216 (2009) |
|
Февраль 2009 |
255 |
1320 |
4475±460 |
Вестник Академии ДНК-генеалогии, т.2 232-251 (2009) |
|
Март 2009 |
98 |
711 |
4700±500 |
J. Genetic Genealogy, 5, 186-216 (2009) |
|
Июнь 2009 |
110 |
804 |
4750±500 |
J. Genetic Genealogy, 5, 186-216 (2009) |
|
Ноябрь 2010 |
148 |
1037 |
4500±470 |
Биохимия, 76, 634-651 (2001) |
|
2023 |
4475±460 |
|||
|
2748 |
4475±470 |
|||
|
Сентябрь 2013 |
647 |
2059 |
4700±480 |
Это последнее положение будет дополнительно
проиллюстрировано ниже.
Теперь, после относительно легкого экскурса в основы ДНК-генеалогии, в последней части настоящей статьи мы остановимся на ДНК-генеалогии несколько более углубленно, описав ее короткую историю, и подведя читателя к возможностям этой области науки. ДНК-генеалогия заявила о себе как о новой научной дисциплине семь лет назад, в 2008 году. Начало было положено обширным анализом в рамках новой науки в то время протяженных (25-маркерных) гаплотипов гаплогруппы R1a Y-хромосомы европейского мужского населения, с расчетом времен жизни общих предков по странам и регионам (результаты были опубликованы в третьем выпуске журнала «Вестник Российской Академии ДНК-генеалогии» в августе 2008 года). Было показано, что мужское население (гаплогруппы R1a) современной Германии имеет общего предка примерно 4600 лет назад, и базовый (предковый) гаплотип его был следующий:
13 25 16 10 11 14 12 12 10 13 11 30 15 9 10 11 11 24 14 20 32 12 15 15 16
25 ноября того же года, то есть через три месяца,
вышла статья Haak et al. (2008), в которой были описаны ископаемые гаплотипы
гаплогруппы R1a, найденные в местечке Эулау, Германия, с датировкой 4600 лет назад.
Ископаемые гаплотипы имели вид
13/14 25 16 11 11 14 X X 10 12/13 X 30 14/15 X X X X X 14 19 X X X X X
(двойные числа в ископаемых гаплотипах – это те,
которые точно определить не смогли, здесь возможны варианты). Знаки X указывают
на пропущенные аллели в ископаемых гаплотипах. Только две аллели (числа в
гаплотипах выше) различались на один шаг, 10 и 11 в четвертом положении
(DYS391), и 19 и 20 ближе к концу гаплотипа (DYS 448). Это вполне естественно,
никто не мог утверждать, что найденные в захоронении останки принадлежали
именно первопредку «германских» R1a, тем более что там была целая семья. Две
мутации разницы по расчетам в рамках ДНК-генеалогии дают всего несколько сотен
лет расстояния между ними, что укладывается в диапазон погрешности расчетов
(Klyosov, 2009a).
Со времени этого первого положительного испытания и демонстрации практического совпадения расчетов и археологических датировок, то есть с лета 2008 года, и начался отсчет новой науки ДНК-генеалогии.
Чтобы понять расчеты и суть
подхода несколько более основательно, рассмотрим, как проводился расчет и на
основании чего. В те времена, 2008-й год, в академической литературе
использовались в основном гаплотипы от 6- до 10-маркерных, были известны
12-маркерный (FTDNA) и 17-маркерный гаплотип (последний был введен в 2006
году), но для хронологических расчетов они практически не использовались. Короткие
же гаплотипы обрабатывались популяционными генетиками с применением
«популяционной» скорости мутации, которую описана в
подробностях в других изданиях. Здесь
отметим только, что она ужасающе неверна, и завышает датировки обыкновенно на
250-350%.
В подходе ДНК-генеалогии
2008 года строилось дерево гаплотипов для проверки на общую симметричность, то
есть на отсутствие выраженных ветвей.
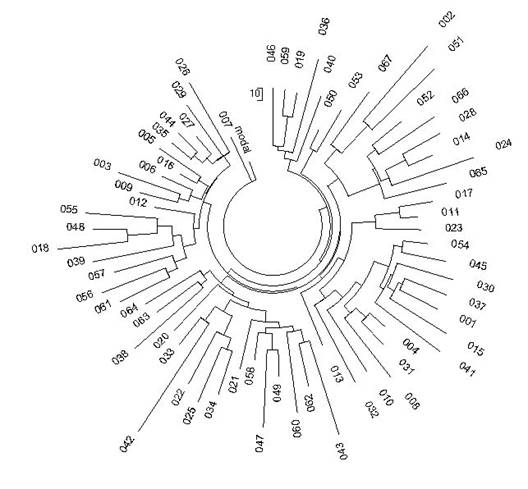
Рис. 2. Дерево из
67 25-маркерных гаплотипов современных немцев гаплогруппы R1a (Клёсов, 2008a).
Дерево выше показывает общую
симметричность (множество мелких ветвей в нем можно анализировать по
отдельности, но опыт показывает, что итоговый результат будет практически такой
же), и его можно количественно обрабатывать как единую систему, то есть что все
67 гаплотипов происходят от одного общего предка. На все 67 гаплотипов в
12-маркерном формате имеется 208 мутаций от приведенного выше базового
гаплотипа, и в 25-маркерном формате - 488 мутаций. В первом случае получаем
208/67/0.02 = 155à183 условных поколений (по
25 лет) от общего предка, то есть 4575±560
лет до общего предка. Во втором случае, для 25-маркерных гаплотипов,
получаем 488/67/0.046 = 158 à187 условных поколений, то
есть 4675±510 лет до общего предка.
Как видим, совпадение почти абсолютное
для расчетов по 12- или 25-маркерным гаплотипам. Здесь (и вообще в
ДНК-генеалогии) на поколение берется 25 лет, это – математическая величина, не
имеющее отношения к продолжительности «бытового поколения», которое есть величина
плавающая, и зависит от обычаев, религиозных верований, эпохи, мора, войн и так
далее. Условное поколение в 25 лет привязано к величине константы скорости
мутации 0.02 (мутаций на условное поколение на 12-маркерный гаплотип), или
0.046 (мутаций на условное поколение на 25-маркерный гаплотип). Константы
скорости мутации для более протяженных гаплотипов
(37-, 67- и 111- маркерных) будут даны позже. Надо сказать, что уже в первом
выпуске Вестника, в июне 2008 года, проводились многочисленные расчеты для 37-
и 67-маркерных гаплотипов, что тогда было неслыханно. Результаты всех этих
расчетов верны до настоящего времени.
Надо
сказать, что неопытный читатель тут же запротестует, что, мол, как можно по
всей Германии, или любом другом регионе судить по каким-то 67 гаплотипам, а там
живут миллионы человек. На это я обычно задаю встречный вопрос – чтобы понять, что море
соленое, нужно ли пробовать воду из всех бухточек? А как же химики делают анализ
растворов, беря пробу всего лишь в долю миллилитра из ведра, цистерны, озера?
Когда делают анализ крови, что, всю кровь из вас выкачивают? Нет. Значит,
вопрос о размере пробы поставлен некорректно. Вопрос на самом деле сводится к
тому, равномерно ли перемешана система, чтобы по малой пробе, или выборке,
судить о всей системе. Этот вопрос в ДНК-генеалогии
давно проработан, и результаты сопоставительных исследований опубликованы
(напр., Клёсов, 2011). В горах – одна ситуация, там в каждой
долине состав гаплотипов (и гаплогрупп/субкладов) может заметно различаться, на
равнине – другая, там гаплотипы более перемешаны за тысячелетия. Но в
любом случае расчеты и выводы по гаплотипам относятся только к той конкретной
выборке, которая рассматривалась, и только сопоставление разных выборок может
показать, насколько система «перемешана», и решить вопрос об обобщении
результатов расчетов и выводов на всю популяцию региона, или только ее
конкретную часть.
Чтобы не быть голословным,
рассмотрим выборку по Германии уже 2014 года. Теперь в ней 286 гаплотипов в 67-
и 65 гаплотипов в 111-маркерном формате (база данных IRAKAZ-2014). Ниже –
дерево 67-маркерных гаплотипов.
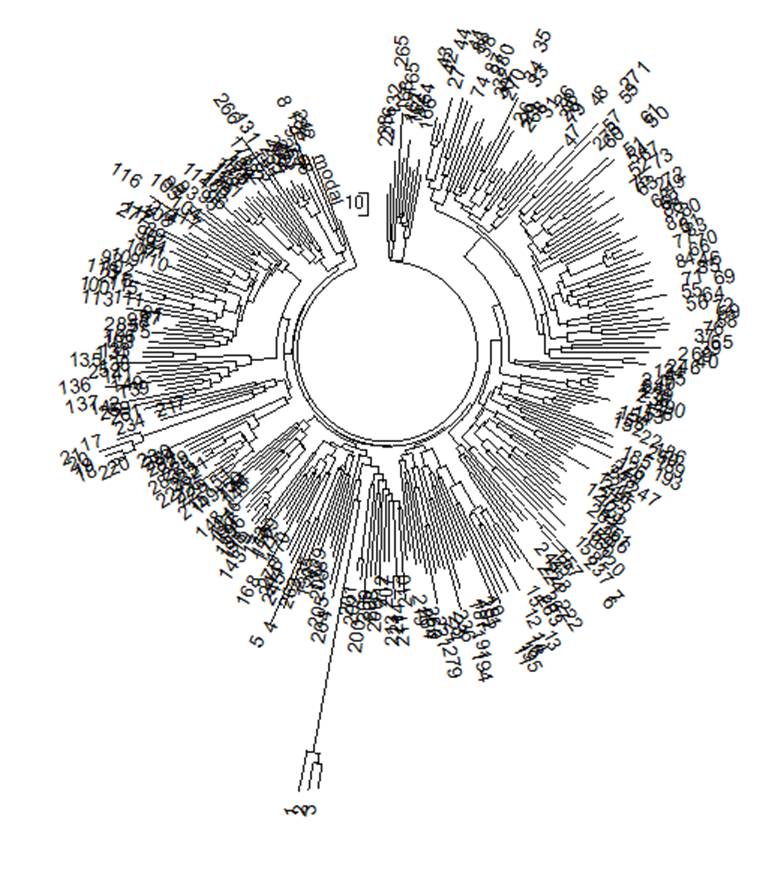
Рис. 3. Дерево из
286 67-маркерных гаплотипов современных немцев гаплогруппы R1a (база данных
IRAKAZ-2014). Три гаплотипа внизу (номера 1, 2, 3) относятся к древнейшему
субкладу гаплогруппы, R1a-M420, и в расчеты (см.
текст) не включались. Расчеты проводили на калькуляторе Килина-Клёсова,
поскольку ручной расчет системы с более чем 22 тысячи аллелей слишком
трудозатратен. Место субклада M420 показано на следующей диаграмме:

Общий предок всех
67-маркерных гаплотипов жил 4848±415 лет назад, всех 67- и 111-маркерных
гаплотипов - 4294±298 лет назад, при округлении получаем 4800±400 и 4300±300 лет
назад. Среднюю величину можно не брать, и так видно, что она практически равна полученным ранее, в 2008 году, и с совершенно другой
выборкой в Германии 4575±560 лет и 4675±510 лет до общего предка,
расчитанным по 12- и 25-маркерным гаплотипам.
Таким образом, обе выборки –
2008 и 2014 гг являются вполне репрезентативными для
целей настоящего исследования.
Помимо того, что в 2008 году
ДНК-генеалогия прошла проверку на реальных (ископаемых) гаплотипах, показав ту
же хронологию, в том же 2008 году, в июне, вышел первый выпуск Вестника,
который позже стал международным. В первом выпуске были две больших статьи –
«Основные правила ДНК-генеалогии» (54 страницы) и «Происхождение евреев с точки
зрения ДНК-генеалогии» (179 страниц). С тех пор вышло 66 выпусков Вестника,
общим объемом более 10 тысяч страниц, все выпуски в открытом доступе (http://aklyosov.home.comcast.net).
C второго выпуска (июль 2008) в разделе «Обращения читателей и персональные
случаи ДНК-генеалогии» публикуются ответы на письма читателей из разных стран
мира. До сегодняшнего выпуска опубликованы персональные истории в виде ответов
на 234 письма читателей. Авторы Вестника – в основном
российские специалисты и любители ДНК-генеалогии, но также специалисты из США,
Италии, Франции, Словении, Сербии, Италии, Германии, Бельгии, Шотландии и
других стран.
С того же
2008 года опубликованы 17 научных статей по ДНК-генеалогии в академических
журналах (все, кроме одной, на английском языке, и одна – в старейшем российском
академическом журнале «Биохимия»), 6 книг по ДНК-генеалогии (5 на русском языке
и одна на сербском, в Белграде), и 82 научно-популярных статьи по
ДНК-генеалогии, из них 55 статей на Переформате – сайте, который выпускается
профессиональными историками. Еще несколько статей на Переформате добавил член
Академии ДНК-генеалогии И.Л. Рожанский, выпускник того же же
Химического факультета МГУ, что и автор настоящей статьи.
Академия ДНК-генеалогии была
создана в том же 2008 году. Как известно, Академия – это сообщество научных единомышленников,
выпускающих свое печатное издание. Пока Академия зарегистрирована только в США,
но в настоящее время готовится к регистрации в России (Москва). Состав членов
Академии не расширяется с 2008 года, и делается это намеренно, чтобы не
«снижать планку» Академии. Два основных требования к членам Академии – знать
свои гаплотип и гаплогруппу, и публиковаться в научных изданиях по теме
ДНК-генеалогии. После регистрации в России состав членов Академии будет
расширяться, и, видимо, значительно, вплоть до десятков и сотен человек.
Научные статьи в области
ДНК-генеалогии публиковались в журналах Human Genetics (2009), J. Gen.
Genealogy (2009), Advances in Anthropology (2011-2014), Биохимия (2011), Eur.
J. Human Genetics (2014), Frontiers in Genetics (2014).
Первой статьей в АА была
наша (совместно с И.Л. Рожанским) статья по константам скоростей мутаций в
гаплотипах Y-хромосомы и их калибровке, то есть количественной связи с
хронологией. Концептуально сделать это было несложно,
поскольку кинетика химических и биологических мутаций – прямая специальность
автора данной статьи, я в свое время заканчивал кафедру академика Н.Н.
Семенова, единственного лауреата Нобелевской премии среди советских химиков, и
написал немало учебников для Высшей школы (в основном издательства Московского
университета и издательства «Наука»), часть из них – с моим учителем, деканом
Химического факультета МГУ чл.-корр. АН СССР И.В. Березиным. За
разработку кинетики химических и биологических реакций я в 1984 году стал
лауреатом Государственной премии СССР по науке и технике, а до того – лауреатом
премии Ленинского комсомола (1978). Так что концептуально разработать количественную
основу ДНК-генеалогии сделать было несложно, но надо было ее верифицировать на тысячах примеров, чтобы она была надежной. Это было сделано
с использованием 3160 гаплотипов (2489 из них – в 67-маркерном формате, из 55 ДНК-генеалогических линий из 8 основных гаплогрупп). Эта
работа была опубликована в Advances in Anthropology в 2011 году, во втором
выпуске журнала после его основания в том же году (Rozhanskii & Klyosov,
2011).
В последующем в том же журнале были опубликованы статьи с детальным описанием гаплогруппы R1a в мире, гаплогруппы R1a в Европе (с описанием 38 ветвей гаплогруппы R1a), гаплогруппы R1b, ДНК-генеалогии и лингвистики, ДНК-генеалогии и древней керамике от трипольской культуры до культур Юго-Восточной Азии и обеих Америк, а также четыре статьи об Африке и происхождении человека с точки зрения ДНК-генеалогии (две в 2012 г, и две – в 2014 г, одна из них редакторская). Одна статья из этих четырех (Klyosov & Rozhanskii, 2012) установила своеобразный рекорд популярности у читателей журнала – по состоянию на 31 декабря 2014 года она была просмотрена 76534 раз, и «скачана» 16273 раз
(http://www.scirp.org/journal/HottestPaper.aspx?JournalID=737).
Так вот, про калибровки. Эти
данные были опубликованы еще в 2011 году (Rozhanskii & Klyosov, 2011). Так
что буду краток, желающие могут прочитать подробности в исходной публикации.
Суть в том, что были взяты генеалогические данные для 13 семей, удовлетворяющие
сформулированным жестким критериям. Эти 13 семей (или «Проектов») были отобраны
из сотен других, которые были менее многочисленны или датировки которых были
менее достоверны, или гаплотипы были короткими, то есть низкого разрешения.
В указанной статье в
Advances in Anthropology приведены многочисленные графики для гаплотипов разной
протяженности, и каждый график иллюстрировал надожность калибровки,
доверительные интервалы и прочее. Статья – редкая по глубине обоснований и
достоверности полученных данных, результаты калибровок выверены на 3160 гаплотипах
из 55 гаплогрупп и субкладов, из них 2489 гаплотипов были 67-маркерными, с которыми
популяционные генетики не только не работают, но не могут и мечтать. Не буду
приводить все калибровочные графики, дам только калибровочную диаграмму для 37-
и 67-маркерных гаплотипов. Здесь на горизонтальной оси – число лет до общего
предка каждой из ДНК-генеалогических «семей», на
вертикальной оси – среднее число мутаций на маркер (поскольку гаплотипы разные
– 37- и 67-маркерные) в гаплотипах, принадлежащих этим семьям.
Рис.
4. Калибровочные взаимоотношения между временем, прошедшим от общих предков
генеалогических серий гаплотипов в каждой группе («генеалогической семье»), и
числом мутаций, накопившхся с того времени, в расчете на маркер в гаплотипах в
37- и 67-маркерном формате. Данные и принцип подхода
опубликованы в журнале Advances in Anthropology (2011, Rozhanskii &
Klyosov) и приведены в дополненном виде И.Л. Рожанским на сайте «Переформат» (
http://pereformat.ru/2014/11/dna-calibration/).
Видно, что есть четкая линейная зависимость между «возрастом» ДНК-линии и числом мутаций на маркер. Так и должно быть, поскольку мутации в гаплотипах (и маркерах) происходят случайным образом и описываются кинетикой первого порядка. Угол наклона корреляционной прямой для 37-маркерных гаплотипов выше, чем у 67-маркерных, поскольку мутации в 37 маркерах (где большая доля «быстрых» по мутациям маркеров) в совокупности происходят чаще, чем в 67-маркерных (где, напротив, маркеры с 38 до 67-го по порядку содержат высокую долю «медленных» маркеров). Отсюда и получились те константы скоростей мутаций, приведенные выше в данной статье: для 37-маркерных гаплотипов 0.00243 мутаций на маркер (0.09 мутаций на гаплотип) за условное поколение, для 67-маркерных гаплотипов 0.00179 мутаций на маркер (0.12 мутаций на гаплотип) за условное поколение. Для 67-маркерных гаплотипов точность калибровки больше, и соответствует погрешности в определении константы скорости мутаций ±2.5 % (И.Л. Рожанский, ссылка приведена выше). Рассмотрение тех нескольких тысяч гаплотипов, упомянутых выше, из 55 гаплогрупп и субкладов (Rozhanskii, Klyosov, 2011) позволило заключить, что использованная нами калибровка и метод расчета дают точность в определении времени жизни предка с точностью ±10 % или меньше в интервале от 500 до 6000 лет назад.
Еще иллюстративный пример. В
Шотландии есть знаменитая «генеалогическая» семья МакДоналдов, их несколько тысяч
человек, на них работает целый штат профессиональных генеалогов, их
документальная генеалогия изучена во всех возможных деталях. И неудивительно,
они берут свое начало от шотландского вождя Сомерледа, среди их предков
наполеоновский маршал МакДоналд и много других выдающихся людей. Основная
группа МакДоналдов гаплогруппы R1a ведет свою линию от Джона Лорда Островов
(John Lord of the Isles), умершего в 1386 году, то есть 628 лет назад. Принимая
25 лет на условное поколение (именно эта величина входит в величины констант
скоростей мутаций, приведенные выше), получаем, что Джон умер 25 условных
поколений назад, то есть жил примерно 26 условных поколений назад. В книге
«Происхождение человека» (2010) я рассмотрел серию гаплотипов потомков Джона,
которая тогда насчитывала сначала (в 2008 году) 68 гаплотипов, затем (в 2010
году) 84 гаплотипа. Среди 68 гаплотипов в 6-маркерном виде было 17 мутаций, и
53 гаплотипа были немутированными, то есть базовыми. Считаем по правилам
ДНК-генеалогии. Сначала по мутациям, используя линейный метод: 17/68/0.0088 = 28±7 условных поколений до Джона
(погрешность велика, поскольку мутаций мало). Теперь считаем без учета мутаций,
используя логарифмический метод: [ln(68/53)]/0.0088 = 28±5 условных поколений назад. 0.0088 – это константа скорости мутаций
для 6-маркерных гаплотипов, см. выше.
Потом появились 60
25-маркерных гаплотипов, в них – 69 мутаций, и из них 18 гаплотипов были
базовыми, то есть идентичными друг другу. Получаем:
Линейный метод – 69/60/0.046
= 25±4 условных поколений.
Логарифмический метод –
[ln(60/18)]/0.046 = 26±7 условных
поколений.
Потом, с расширением тестирования, появились 84 25-маркерных гаплотипов, и в них во всех содержалось 109 мутаций. В тех же 84 гаплотипах в 12-маркерном формате было 44 мутации. Получаем:
Для 12-маркерных
гаплотипов 44/84/0.02 = 26±5 условных поколений.
Для 25-маркерных гаплотипов
109/84/0.046 = 28±4 условных
поколений.
Здесь 0.02 и 0.046 – это константы скорости мутации
для 12- и 25-маркерных гаплотипов, соответственно.
Итак, наблюдается совершенно разумное совпадение с документальной генеалогией по Джону и его потомкам: документальная генеалогия дает 628 лет до годовщины смерти Джона (1386 год), а ДНК-генеалогия дает 650-700 лет до времени его жизни (1314-1364 год), то есть практически абсолютное совпадение в контексте данного исследования, учитывая, что год его рождения неизвестен.
И таких примеров десятки, если не сотни, когда ДНК-генеалогия дает либо совпадения с документальной генеалогией (в пределах погрешности расчетов), либо дополняет документальную генеалогию, когда ее данные фрагментарные или вовсе отсутствуют, кроме приблизительных датировок.
Продолжим сравнительное рассмотрение ДНК-генеалогии и
популяционной генетики, их расчетные методы и характер выводов. Рассмотрим
пример, имеющий важное историческое значение. Он имеет прямое отношение к
легендарным ариям, которые по историческим сведениям (скорее, предположениям)
перешли в Индию примерно 3500 лет назад. Это впервые было высказано, видимо,
немецко-английским ученым-ориенталистом Максом Мюллером (1823-1900), и эта
дата приводится в книге Гордона Чайлдса
«Арии. Основатели европейской цивилизации» (1926). Он пишет: «...Ригведа была создана вскоре после 1400 года до н.э., и вторжение ариев в Индию
относится примерно к тому же времени» (то есть примерно 3400 лет назад). Но
в следующем абзаце Чайлдс продолжает – «Недавно
вызов этой традиционной точке зрения был брошен сразу с двух сторон. Паргитер считает,
что проникновение ариев в Индию началось задолго до составления ведических
гимнов... Изучение генеалогий правителей привело упомянутого автора к заключению,
что арии... вторглись в Индию скорее
ближе к началу, чем к концу 2-го тысячелетия до н.э.
... Однако в последние годы она была
оспорена и с другой стороны. Брюннхофер и некоторые другие исследователи утверждают,
что ... проникновение ариев в Индию следует относить к гораздо более позднему
времени, чем это обычно предполагается».
Итак, имеем датировки для перехода ариев в Индию – примерно 3400 лет назад, примерно 4000 лет назад, и позже, скажем, 3000-2800 лет назад. Такой разброс и остался с 1926 года до настоящего времени. Поскольку четкой археологии, которая дала бы уточнение датировок, нет, то историческая наука так и осталась в отношении ариев в Индии на том же уровне, что и 90 лет назад. Индийский эпос в основном иносказательный, из него для исторической науки много не получить. Древние названия местностей в Индии, как Арьяварта, датировкам не помогают.
Давайте посмотрим на эти данные, и сравним, что
говорит об ариях в Индии ДНК-генеалогия. В сети есть Проект Индии (FTDNA), это
фактически база данных индийских гаплотипов. В ней 187 гаплотипов в
37-маркерном формате, из них 64 гаплотипа гаплогруппы R1a, то есть 34% от всех.
Дерево всех 187 гаплотипов выглядит следующим образом.
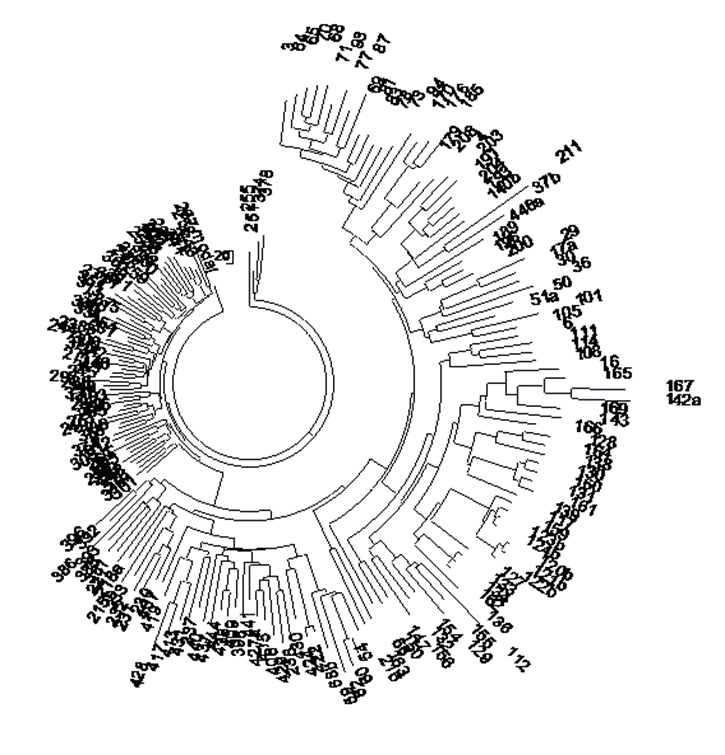
Рис.
5. Дерево 187 индийских гаплотипов Y-хромосомы в
37-маркерном формате. Слева – плотная ветвь из 64 гаплотипов
гаплогруппы/субклада R1a-L657, с общим предком, жившим 4750±500 лет назад;
справа и внизу – серии древних индийских гаплотипов гаплогрупп (по часовой
стрелке) Н, L, Q, L, J-M304, O, C, L, C, J1-M267, J2b-L282, J2-M172, R2-M124, Q
(повторы гаплогрупп – это отдельные ветви этих гаплогрупп). Общие предки ветвей
справа и внизу жили 8-12 тысяч лет назад. Построено по данным Индийского проекта
FTDNA (
https://www.familytreedna.com/public/India/default.aspx?section=yresults)
Слева – все 64 гаплотипа гаплогруппы R1a, которые образовали плотную ветвь, составленную из субклада R1a-L657. То, что ветвь вполне однородная, показывает следующее дерево, уже только из гаплотипов данной ветви R1a-L657 (рис. 6). Однородность дерева уже показывает, что все гаплотипы родственные, все происходят от одного общего предка, во всяком случае с той точностью, которая нас в данном случае вполне устраивает. Посчитаем, когда этот предок жил. Во всех 64 гаплотипах в 37-маркерном формате – 924 мутации, что дает 924/64/0.09 = 160 à 190 условных поколений, то есть 4750±500 лет назад. Расчет с помощью калькулятора Килина-Клёсова (Килин и Клёсов, 2014; Клёсов, 2014a, b), основанного совершенно на другом принципе расчета, а именно по каждому маркеру отдельно, используя калиброванные абсолютные константы скорости мутаций для каждого из 37 маркеров, дал время жизни общего предка гаплотипов R1a в Индии 4964±548 лет назад, то есть, округляя, получаем 5000±550 лет назад. Это та же величина, что и полученная «линейным методом» 4750±500 лет назад, в пределах погрешности расчетов.
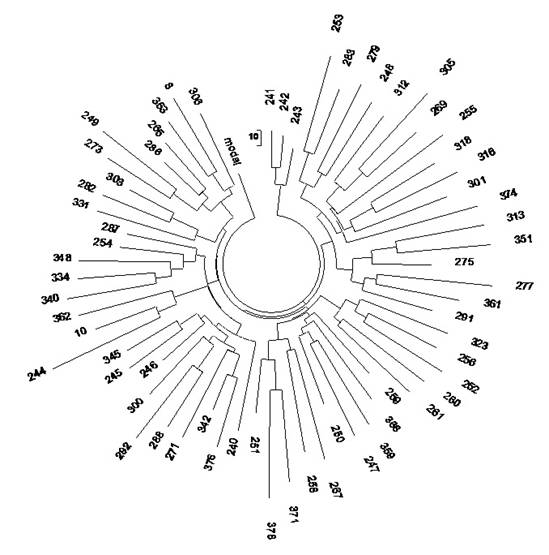
Рис.
6. Дерево 64 индийских гаплотипов гаплогруппы R1a-L657
в 37-маркерном формате. Общий предок дерева жил 4750±500 лет назад. Построено
по данным Индийского проекта FTDNA (
https://www.familytreedna.com/public/India/default.aspx?section=yresults)
Но это - время, отдаляющее нас от времени жизни общего предка гаплотипов группы R1a, живущих в настоящее время в Индии. Это вовсе не значит, что этот предок жил в Индии 4750-5000 лет назад. В Индию его потомки пришли примерно 3400-4000 лет назад, то есть на тысячу лет позже времени жизни своего общего предка. Где же этот предок жил, откуда в Индию пришли арии?
Чтобы ответить на этот
вопрос, рассмотрим базовый гаплотип индийских R1a. Он
– следующий (к нему, к временам примерно 5000 лет назад, сходятся все 64
указанные гаплотипа):
13 25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
А вот – предковый гаплотип этнических русских гаплогруппы R1a, общий предок которых жил на Русской равнине 4900 лет назад (гаплотип приведен в книгах «Происхождение славян» (А.А. Клёсов, стр. 26, 2013), и «Арийские народы на просторах Евразии» (А.А. Клёсов, К.А. Пензев, стр. 246, 2014):
13 25 16 11 11 14 12 12 10 13 11 17 – 15 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 16 16 18 19 35 38 14 11
Мы видим, что предковые
гаплотипы ариев и этнических русских очень похожи, между ними на вид всего 5
мутаций. На самом деле там всего 3.5 мутаций, потому что все отличающиеся
аллели дробные. 3.5 мутаций между двумя 37-маркерными базовыми гаплотипами
разделяют эти гаплотипы (то есть предков индийцев и этнических русских) на
3.5/0.09 = 39 à 41 условных поколений, или
примерно 1025 лет. То есть их общий предок (субклад R1a-Z645, см. диаграмму
ниже) жил примерно за 500 лет до появления общих предков сегодняшних этнических
русских гаплогруппы R1a и индийцев той же гаплогруппы, или примерно 5500 лет
назад.
• R1a1a1 M417
• • R1a1a1b Z645
• • • R1a1a1b1 Z283
• • • • R1a1a1b1a Z282
• • • • • R1a1a1b1a2 Z280
• • • R1a1a1b2 Z93
• • • • R1a1a1b2a Z94, L342.2
• • • • • R1a1a1b2a1 L657
Примерно ту же датировку
дают снипы (то есть SNP, необратимые мутации в Y-хромосоме). По данным
Владимира Таганкина, который серьезно занимается снипами Y-хромосомы, между
Z645 и Z283 всего два снипа, между Z283 и Z282 всего один снип, и между Z282 и
Z280 – всего три снипа. «Всего» - потому что по
предварительным данных один снип в среднем образуется каждые 100-150 лет назад,
данные еще недостаточно откалиброваны. Поэтому при шести снипах между
Z645 и Z280 (последний образовался примерно 5000 лет назад), Z645 (общий предок
ариев и этнических русских) образовался примерно 5600-5900 лет назад. Они
действительно близкие родственники, в пределах нескольких столетий. Более того,
при наличии древней арийской топонимики на Русском севере становится ясным, что
арии вовсе не замыкались в южных степях, как полагают некоторые археологи на
основе материальных признаков степной и лесостепной полосы юга России и
Украины. Ясно, что арии заселяли всю Русскую равнину с юга до севера, имели
практически такие же гаплотипы, как и предки современных этнических русских (с
общим предком всего за несколько веков до тех времен). Поэтому совершенно разумно считать носителей гаплогруппы R1a-Z645
ариями, как и их потомков, носителей субкладов Z93-Z94-L657 и Z283-Z282-Z280.
Они все современники друг друга.
Похоже, что в Индии осталось
небольшое количество особенно древних носителей гаплогруппы R1a, и они иногда выявляются
в джунглях, но систематических исследований их не проводилось, субклады их не
определялись, в высших кастах (и в кастах вообще) они
не представлены, они попадают при классификации или в низшие касты (lower
caste), или в племена (tribes). Видимо, они попали в Индию в ходе древнейших
миграций из Южной Сибири на запад, когда носители R1a в итоге прибыли в Европу
примерно 9-10 тысяч лет назад (Klyosov, 2009b, 2012). Но эти данные получены на
коротких гаплотипах, в которых ошибки тестирования проявляются намного больше,
чем при работе с протяженными гаплотипами. А поскольку
данные по этим древним R1a в Индии фрагментарные (в известном списке Sengupta
[Sengupta et al., 2006] их, например, практически нет, за исключением, быть
может, гаплотипов 4, 95, 109 на дереве гаплотипов на рис. 7), то
систематического изучения их пока не проводилось, субклады неизвестны.
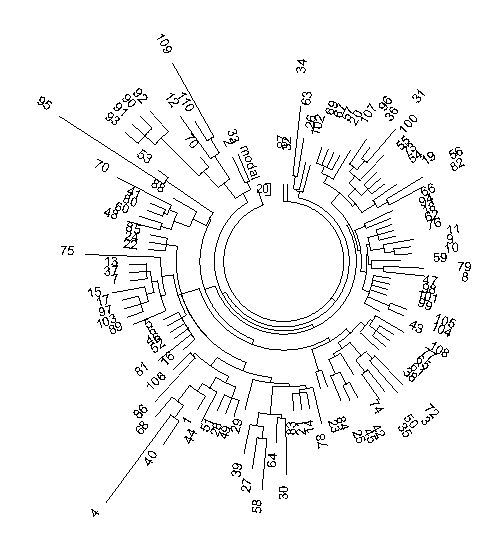
Рис. 7. Дерево из 110
10-маркерных гаплотипов гаплогруппы R1a в индийских кастах и племенах.
Построено по данным Sengupta et al. (2006). Формат гаплотипов
- DYS 393, 390, 19, 391, 388, 439, 389-1, 393, 389-2, 461).
На данных Sengupta et al. (2006) стоит остановиться чуть подробнее, поскольку именно «расчеты» этих самых известных данных по Индии (более тысячи 10-маркерных гаплотипов в выборке, из них 110 гаплотипов гаплогруппы R1a) приводят популяционных генетиков к тому, что общий предок носителей R1a в Индии жил более 14 тысяч лет назад Дерево гаплотипов на рис. 7 не слишком симметричное, но считать вполне можно, на что указывает и полученный базовый гаплотип дерева (здесь приведены 9 маркеров, поскольку десятый, DYS461, не входит в список 37-маркерных гаплотипов, которые будут приведены ниже для сравнения, Х – маркеры, которые не определяли в работе Sengupta)
13 25 15 10 Х Х Х 12 10 13 11 17
Все 110 гаплотипов содержат 344 мутации, что дает 344/110/0.018 = 174 à 211 условных поколений, то есть 5275±600 лет до общего предка. Здесь 0.018 – константа скорости мутации для 9-маркерного гаплотипа, равная сумме констант для всех 9 маркеров, определенных еще в 2006 году Чандлером (Chandler, 2006).
Сравним полученный базовый
гаплотип с таковым для индийцев субклада R1a-L657, приведенным выше в
37-маркерном формате (общий предок жил 4750±500 лет назад):
13 25 16 10 11 14 12 12 10 13 11 17 – 16 9 10 11 11 24 14 20 32 12 15 15 16 – 11 12 19 23 15 16 18 19 35 39 13 11
Если переписать его в виде, удобном для сравнения
13 25 16 10 Х Х Х 12 10 13 11 18
то мы увидим, что базовый гаплотип, полученный по
данным Sengupta et al. (2006) (включающих низшие и высшие касты и племена), и
базовый гаплотип, полученный по данным индийского проекта FTDNA, различаются
всего на одну мутацию (выделено), а на самом деле всего на 0.55 мутаций, если
усреднить по всем аллелям маркера DYS19. Другими словами, это одинаковые
предковые гаплотипы, и одинаковая датировка, в пределах погрешности расчетов,
безотносительно, 9-маркерные гаплотипы или 37-маркерные. Некоторое завышение
датировки по данным Sengupta (2006), которое, впрочем, укладывается в
погрешности, вызвано тремя особенно мутированными гаплотипами (под номерами 4,
95, 109, см. рис.7), на которые приходится 19 мутаций, и при их снятии получаем
325/107/0.018 = 169 à 203 условных поколений, или
5075±580 лет до общего предка.
Итак, становится понятным смысл нового термина – «молекулярная история», т.е. создание исторических реконструкций исходя из молекулярных характеристик ДНК потомков, а также ископаемых предков. Поскольку далекие предки, передвигаясь, несли в новые края языки, то, прослеживая миграции предков, происходившие сотни, тысячи и десятки тысяч лет назад, можно получать сведения о миграции языков во времена столь глубокой древности. Сопоставление этих реконструкций с данными лингвистики, полученными принципиально другими методами, может позволить получать более обоснованные сведения в области языкознания, проверять существующие концепции и приходить к новым, совершенно неожиданным концепциям и идеям. Так биохимия, физическая химия, химическая кинетика повернулись своими неожиданными и непредсказуемыми ранее гранями к наукам гуманитарным, историческим, лингвистическим.
Литература
Интернет-сайт
«Sorenson Molecular Genealogy Foundation»,
http://www.smgf.org/ychromosome/marker_details.jspx?marker/
Интернет-сайт R1a,
http://r1a.org/irakaz-v03.xls, http://r1a.org/
Интернет-сайт «A
Free Public Service from Family Tree DNA»,
http://www.ysearch.org/,
http://www.smgf.org/pages/ydatabase.jspx/
http://www.familytreedna.com/public/R1aY-Haplogroup/default.aspx?section=yresults,
http://www.familytreedna.com/public/R1a/default.aspx?section=yresults
Килин, В.В., Клёсов, А.А. (2014) Принципиально новый
калькулятор расчета времен до общих предков (ТМRCA)
серий гаплотипов во всем диапазоне от сотен до миллионов лет назад, основанный
на модели случайных блужданий. Вестник Академии ДНК-генеалогии, т. 7, № 3,
438-478.
Клёсов, А.А. (2008а) Откуда появились славяне и
«индоевропейцы»? Ответ дает ДНК-генеалогия. Вестник Российской Академии
ДНК-генеалогии, т. 1, №3, 400-477.
Клёсов, А.А. (2008b) Руководство к расчету
времен до общего предка гаплотипов Y-хромосомы и таблица возвратных мутаций.
Вестник Росийской Академии ДНК-генеалогии, т. 1, № 5, с. 812-835.
Клёсов, A.A. (2011) Биологическая химия как основа ДНК-генеалогии и зарождение «молекулярной
истории». Биохимия, 76, № 5,
636 – 653.
Клёсов, А.А. (2013a) Происхождение славян. М.,
Алгоритм, 511 с.
Клёсов, А.А. (2013b) Занимательная ДНК-генеалогия.
М., Вече, 168 с.
Клёсов, А.А. (2014a) Опыт работы с ККК
(калькулятором Килина-Клёсова) расчета времен до общих предков (ТМRCA), основанный на модели
случайных блужданий и с использвоанием 111 индивидуальных констант скоростей
мутаций. Часть 1. Вестник Академии ДНК-генеалогии, т. 7, № 4, 626-638.
Клёсов, А.А. (2014b) Опыт работы с ККК
(калькулятором Килина-Клёсова) расчета времен до общих предков (ТМRCA), основанный на модели
случайных блужданий и с использованием 111 индивидуальных констант скоростей
мутаций. Часть 2. Вестник Академии ДНК-генеалогии, т. 7, № 5, 758-769.
Клёсов, А.А., Пензев, К.А. (2014) Арийские народы на
просторах Евразии (М, изд. «Книжный мир»). Глава 7, стр. 223-234.
Chandler,
J. F. (2006) Estimating per-locus mutation rates. J. Genet. Geneal.,
No. 2, 27-33.
Elhaik, E.,
Tatarinova, T.V., Klyosov, A.A., Graur, D. (2014) The 'extremely ancient' chromosome that isn't: a
forensic bioinformatic investigation of Albert Perry's X-degenerate portion of
the Y chromosome". Eur. J. Human Genetics (22 January 2014) doi:10.1038/ejhg.2013.303
(2014)
Haak, W., Brandt, G., de Jong, H.N., Meyer, C.,
Ganslmeier, R., Heyd, V.,
Hawkesworth, C., Pike, A.W.G., Meller, H., Alta, K.W. (2008) Ancient DNA,
Strontium isotopes, and osteological analyses shed light on social and kinship organization
of the Later Stone Age. Proc. Natl. Acad. Sci. US., 105, 18226–18231.
Klyosov,
A.A. (2008) Basic rules of DNA genealogy (Y-chromosome). Mutation rates and their calibration. Proceedings
of the Russian Academy of DNA Genealogy, 1, No. 1, 3-53.
Klyosov,
A.A. (2009a) DNA Genealogy, mutation rates, and some historical evidences
written in Y-chromosome. I. Basic principles and the
method. J. Genetic Genealogy, 5, 186-216.
Klyosov,
A.A. (2009b) DNA Genealogy, mutation rates, and some historical evidences
written in Y-chromosome. II. Walking the map. J.
Genetic Genealogy, 5, 217-256.
Klyosov, A.A. (2009c) A
comment on the paper: Extended Y chromosome haplotypes resolve multiple and unique lineages
of the Jewish Priesthood. Human Genetics, 126, 719-724.
Klyosov, A.A. (2012)
Ancient history of the Arbins, bearers of haplogroup R1b, from Central Asia to
Europe, 16,000 to 1500 years before present. Advances in Anthropology, 2, No.
2, 87-105.
Klyosov,
A.A., Rozhanskii, I.L. (2012a) Haplogroup R1a as the Proto Indo-Europeans and
the legendary Aryans as witnessed by the DNA of their current descendants. Advances in Anthropology, 2, No. 1, 1-13.
Klyosov,
A.A., Rozhanskii, I.L. (2012b) Re-examining the “Out of Africa” theory and the
origin of Europeoids (Caucasoids) in light of DNA genealogy. Advances in
Anthropology, 2, No. 2, 80-86.
Klyosov,
A.A., Rozhanskii, I.L., Ryanbchenko, L.E. (2012) Re-examining the Out-of-
Africa theory and the origin of Europeoids (Caucasoids). Part
2. SNPs, haplogroups and haplotypes in the Y
charomosome of Chimpanzee and Humans. Advances in Anthropology, 2, No. 4, 198-213.
Klyosov,
A.A., Mironova, E.A. (2013) A DNA genealogy solution
to the puzzle of ancient look-alike ceramics across the world. Advances in
Anthropology, 3, No. 3, 164-172.
Klyosov,
A.A. (2014) Reconsideration of the "Out of
Africa" concept as not having enough proof. Advances in Anthropology, 4,
No. 1, 18-37.
Klyosov, A.A., Tomezzoli, G.T. (2013) DNA genealogy and linguistics. Ancient Europe. Advances in Anthropology, 3,
No. 2, 101-111.
Rozhanskii,
I.L., Klyosov, A.A. (2011) Mutation rate constants in DNA genealogy (Y chromosome).
Advances in Anthropology, 1, No. 2, 26-34.
Rozhanskii, I.L., Klyosov, A.A. (2012) Haplogroup R1a, its subclades and
branches in Europe during the last 9,000 years.
Advances in Anthropology, 2, No. 3, 139-156.
Sengupta, S., Zhivotovsky, L.A., King, R., Mehdi, S.Q.,
Edmonds, C.A., Chow, C.-E.T., Lin, A.A., Mitra, M., Sil, S.K., Ramesh, A., Rani,
M.V.U., Thakur, C.M., Cavalli-Sforza, L.L., Majumder, P.P., Underhill, P.A.
(2006) Polarity and temporality of high-resolution Y-chromosome distributions
in India identify both
indigenous and exogenous expansions and reveal minor genetic influence of
Central Asian pastoralists. Am. J. Hum. Genet. 78,
202–221.
Sharma,
S., Rai, E., Sharma, P., Jena, M., Singh, S., Darvishi, K., Bhat, A.K.,
Bhanwer, AJS, Tiwari, P.K., Bamezai,
R.N.K. (2009) The Indian origin of paternal haplogroup R1a1* substantiates the
autochthonous origin of Brahmins and the caste system. J. Human Genetics, 54,
47–55.
DNA GENEALOGY AS
MOLECULAR HISTORY
Anatole
A. Klyosov
This article presents the basis of DNA genealogy, a new field of
science, which is currently emerging as an unusual blend of biochemistry,
history, and chemical kinetics, and shows its principal difference from
population genetics. The methodology of the new approach is comprised of
chemical, or biological kinetics applied to a pattern of mutations in certain non-recombinant
fragments of Y chromosome. The goal of the analysis is to translate DNA mutation
patterns into time spans to the most recent common ancestors of a given
population, or tribe, and to the dating of ancient migration routes. To
illustrate this approach, time spans to the ancient common ancestors are
calculated for ethnic Kyrgyz of haplogroup R1a (the major haplogroup among
Kyrgyz), as well of the present day Germans and Hindu of the same haplogroup.
The common ancestors of two identified groups of the present day Kyrgyz-R1a
according to the pattern of their mutations in the DNA lived 1325±290 and
775±450 years before present (the first group dominates among Kyrgyz, based on
a rather limited albeit random selection of tested people), with their common ancestor
who lived around 1650 years before present, that is approximately in the 4th
century. He belonged the most likely to Scythians or Sarmats, though those
hypotheses should be examined and verified with historians, and either contribute new knowledge to historical studies, or,
conversely, the hypotheses should be reconsidered and corrected. A common
ancestor of present day Germans of haplogroup R1a lived, based on various
random selections of haplotypes sampled in 2008 and 2014, and based on calculations
employing different haplotype format, that is 12-, 25-, 67-, and 111-marker
haplotypes, 4800±400, 4300±300, 4575±560, and 4675±510 years before present.
The figures indicate a rather common range of margins of error in DNA
genealogy. The figures also show that the sampling in both 2008 and 2014 was
rather representative, since it is a key question in DNA genealogy, namely,
what is a justification of using limited selections of dozens and hundreds of
haplotypes and calculating timespans to their common ancestors, when the populations
themselves are numbered in millions. In this particular case the results of the
DNA genealogy calculations were later confirmed by the archaeological data
(Haak et al., 2008), according to which the earliest excavated bearers of
haplogroup R1a in Germany lived 4600 years before present. A common ancestor of
present day Indians (Hindu) of haplogroup R1a, which are largely represented in
Indian upper castes (up to 72%), lived 4750±500 years ago, which within the
margin of error equal to that of the ethnic Russians of haplogroup R1a
(4900±500 years ago), and their base (ancestral) haplotypes are almost
identical (unlike the ancestral haplotypes of predominantly R1b haplogroup in
Western and Central Europe, which are practically absent in India). It is
suggested that the approach described in this paper lays the foundation for
“molecular history”, in which the principal tool is high-technology analysis of
DNA molecules of both our contemporaries and excavated ancient DNA samples.
Key words: DNA genealogy, haplotype, haplogroup, mutations, Y-chromosome
http://aklyosov.home.comcast.net
Академия
ДНК-генеалогии
Бостон, Массачусеттс
http://aklyosov.home.comcast.net
The
Academy of DNA Genealogy
Boston,
Massachusetts, USA