Электронный
математический и медико-биологический журнал. - Т. 14. -
Вып. 1. - 2015. - URL:
http://www.smolensk.ru/user/sgma/MMORPH/TITL.HTM
http://www.smolensk.ru/user/sgma/MMORPH/N-45-html/TITL-45.htm
http://www.smolensk.ru/user/sgma/MMORPH/N-45-html/cont.htm
УДК 57.017.64
О росте живых
организмов
Ó
2015 г. Седова
Г. П.
В данной статье
я имею намерение подвести итог своим исследованиям по количественному росту
живых организмов, которые правильнее
отнести к прикладной математике. Конспективно изложу
выводы из своих статей,
посвященных данной теме,
и постараюсь показать тот
путь, который меня
к ним привел.
Ключевые слова: количественный рост живых организмов.
Если говорить математическим языком, то итог моих
исследований сводится к выводу, что рост всех биологических объектов, под которыми
понимаются как целые организмы, так и отдельные ткани и органы, происходит по
простой формуле:
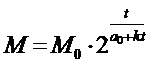 (1)
(1)
где M0 -
начальная масса, t - возраст объекта, a0 - начальный
темп клеточных делений,
k - коэффициент замедления
роста.
В сущности, эта формула представляет собой
аналитическое выражение кривой роста M=f(t). Формула эта эмпирическая, основанная на математическом
анализе экспериментальных данных. В физике таких формул много.
Но я сразу должна признать, что практического значения
эта формула не имеет, т.к. содержит параметры, a0 и k, биологический
смысл которых нам неизвестен. Начальный темп клеточных делений, коэффициент
замедления роста – это чисто формальные, математические определения, а что в
материальном отношении скрывается за этими параметрами, мы не знаем. От математики
нельзя здесь ждать какой-либо помощи, т.к. это, хотя и точная, но абстрактная
наука, имеющая дело только с числами, с величинами, и выявляющая между ними существующие
соотношения.
Биологический
смысл двух параметров a0 и k скрыт где-то в глубине живой материи. Наши
биологические знания не позволяют до него добраться, мы не в состоянии изменять эти параметры. Ну а если
позволить себе пофантазировать, и представить на минутку, что мы это можем, то
изменяя параметр k, мы могли бы изменять по своему усмотрению предельные
размеры организмов, а меняя параметр a0, мы могли бы изменять время достижения организмом предельного
размера, а оно коррелирует с продолжительностью жизни
организма. Верующие в Бога люди скажут, что Бог сознательно ограничил наши
возможности. Ну а как материалистически мыслящий человек, я считаю, что
биология находится еще на низком, в основном описательном уровне развития, она
далека от точных наук. Мы, в сущности, ничего не знаем о росте, ничего не знаем
о митозе, этом главном процессе, лежащем в основе роста. Все стадии митоза подробно
описаны с картинками, но мы смотрим на
эти картинки, как дети, и не можем понять, почему это хромосомы сначала
собираются в экваториальной плоскости, а затем расходятся к полюсам, какие силы заставляют их так себя вести. Не
можем ответить на вопрос: что заставляет зиготу дробиться, а затем бластомеры
расти. Хотя формула (1) не представляет практического интереса ввиду ограниченности
наших знаний по росту живых организмов, но теоретический интерес она представляет,
т.к. получена на основании анализа экспериментальных данных. По этой причине я
остановлюсь на ее выводе и постараюсь изложить это более понятным языком, чем
сделано в моих статьях.
Опорным пунктом для вывода этой формулы является общеизвестный,
доказанный экспериментаторами факт, состоящий в том, что популяция одноклеточных
организмов, растущих независимо друг от друга в данной питательной среде, при
наличии всех необходимых для жизни условий, удваивает свою массу с одинаковым
периодом. Математически это можно выразить следующей формулой:
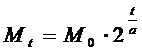 , (2)
, (2)
где Mt - масса популяции в момент времени t, M0 - начальная масса, t - время, a - период удвоения массы.
Здесь a - величина постоянная для каждого вида одноклеточных
организмов, a = const . Поставим
перед собой задачу: распространить эту формулу на многоклеточный организм. Ведь
многоклеточный организм это, в сущности, тоже популяция, но популяция связанных
между собой клеток. Что может измениться в этой формуле в результате этой связи?
Начальная масса в обоих случаях должна присутствовать, время - независимая от
нас величина. Следовательно, если что изменится в формуле (2), так это период
удвоения a. Из повседневных наблюдений мы
знаем, что с возрастом рост многоклеточного организма замедляется, имеется
ввиду замедление не в абсолютных единицах массы, а в относительных. Значит,
для многоклеточного организма в формуле (2) вместо постоянной величины a нужно
поставить некоторую функцию от возраста, т.е. величину a(t). Формула (2) в применении к многоклеточному организму
примет вид:
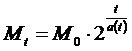 (3)
(3)
Здесь a(t) - неизвестная
нам функция от возраста организма. Нам заранее неизвестно, какой вид она имеет.
Более того, может оказаться, что эта зависимость a(t) не может
быть выражена четкой формулой, а имеет вид корреляции, т.е. зависимость
прослеживается, но осложнена другими факторами.
Для того, чтобы выяснить вид
функции a(t) обратимся
к экспериментальным данным по росту разных организмов. Возьмем, например, данные
по эмбриональному росту человека. Экспериментаторы составили таблицу изменения
массы эмбриона человека от возраста. Она имеет следующий вид:
Возраст эмбриона Масса эмбриона
t1 ………………………………
M1
t2………………………………..M2
t3
……………………………….M3
…………………………………….
…………………………………….
tn...........................................Mn
Из формулы (3) получим общее выражение для величины a(t):
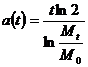 .
.
Подставим
в это общее выражение конкретные числовые значения, получим
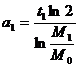
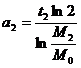
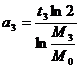 ………
………
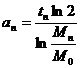 .
.
Мы
получили последовательность чисел a1,a2 , a3 ,……..an
. Нанесем ее на график a=f(t).
Конкретный
график для эмбрионального роста человека, построенный на основании экспериментальных
данных, взятых из книги И. И. Шмальгаузена «Рост и дифференцировка» имеет
следующий вид.
|
Возраст, |
Масса, |
Период |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сутки |
г |
удвоения |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
массы, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
сутки |
|
|
|
||||||||||
|
12 |
0,0002 |
0,785 |
|
|
|||||||||||
|
16 |
0,0034 |
0,826 |
|
|
|||||||||||
|
20 |
0,047 |
0,863 |
|
|
|||||||||||
|
30 |
0,72 |
1,107 |
|
|
|||||||||||
|
56 |
4 |
1,894 |
|
|
|||||||||||
|
84 |
20 |
2,633 |
|
|
|||||||||||
|
112 |
120 |
3,248 |
|
|
|||||||||||
|
140 |
285 |
3,918 |
|
|
|||||||||||
|
168 |
635 |
4,555 |
|
|
|||||||||||
|
196 |
1220 |
5,181 |
|
|
|||||||||||
|
204 |
1617 |
5,336 |
|
|
|||||||||||
|
210 |
1868 |
5,463 |
|
|
|||||||||||
|
218 |
1990 |
5,657 |
|
|
|||||||||||
|
224 |
2107 |
5,801 |
|
|
|||||||||||
|
232 |
2310 |
5,987 |
|
|
|||||||||||
|
238 |
2424 |
6,131 |
|
|
|||||||||||
|
246 |
2765 |
6,306 |
|
|
|||||||||||
|
252 |
2806 |
6,457 |
|
|
|||||||||||
|
253 |
2820 |
6,481 |
|
|
|||||||||||
|
259 |
2878 |
6,63 |
|
|
|||||||||||
|
260 |
2900 |
6,654 |
|
|
|||||||||||
|
266 |
3016 |
6,8 |
|
|
|||||||||||
|
267 |
3030 |
6,822 |
|
|
|||||||||||
|
273 |
3186 |
6,962 |
|
|
|||||||||||
|
274 |
3200 |
6,986 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
280 |
3321 |
7,13 |
|
|
|
|
|
|
|
|
|
|
|
|
|
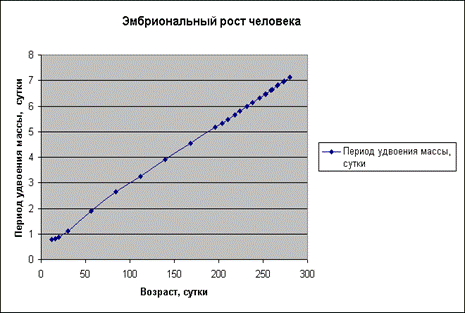
Экспериментальные
данные взяты из
книги И. И. Шмальгаузена «Рост и дифференцировка» ,
т.1, с. 75 .
Мы видим, что график a=f(t) представляет собой прямую линию.
Выполнив подобные вычисления, и построив
соответствующие графики для других организмов, по которым имеются экспериментальные
данные по росту, мы получим тот же результат.
Прямолинейный характер зависимости имеет место как для эмбрионального, так и для постэмбрионального
роста.
За M0 в случае эмбрионального роста можно взять массу
зиготы, примерно равную ![]() г, или наименьшую массу, которую
можно измерить. Для постэмбрионального роста за M0 возьмем массу в момент рождения.
г, или наименьшую массу, которую
можно измерить. Для постэмбрионального роста за M0 возьмем массу в момент рождения.
Необходимо иметь ввиду, что
экспериментальные данные должны быть выполнены с достаточной точностью. Никакие приближенные данные, взятые из источников, где не требуется
большая точность, например, из журналов зоотехнического, экологического направлений,
для данного анализа непригодны.
Линейный характер указанной зависимости имеет место не
только для целых организмов, но и для отдельных органов. Здесь можно сослаться
на эксперименты акад. И. И. Шмальгаузена по измерению роста органов куриных
эмбрионов. Зависимость a=f(t), построенная на основании этих измерений, также
представляет собой прямую линию.
Поставим вопрос: Если мы будем рассматривать не
отдельно эмбриональный и постэмбриональный рост многоклеточного организма, а
совместно, отсчитывая возраст от зиготы до достижения предельного размера, изменится
ли при этом линейный характер зависимости a=f(t)? Математика показывает: нет, не изменится. Получается,
что рождение организма и связанное с ним коренное изменение условий существования,
не искажает линейный характер указанной зависимости. Причина ее заложена
глубже.
Итак, мы приходим к следующему выводу: когда клетки
(одноклеточные организмы) растут независимо друг от друга, они сохраняют
постоянный темп клеточных делений. Их клеточный цикл по времени не меняется на
протяжении большого количества поколений (исчисляемого тысячами, а у некоторых
и более). Причиной замедления роста здесь могут быть только факторы внешнего
порядка, например, истощение питательных веществ, ограничение жизненного
пространства, изменение температуры,
влажности и других
условий существования.
Как только клетки объединились в многоклеточный
организм, их клеточный цикл закономерно удлиняется с возрастом даже при наличии
всех необходимых для роста условий. Это удлинение
разное для разных биологических объектов и определяется параметрами a0 и k. В этом случае причиной неуклонного замедления роста,
как отмечал акад. И. И. Шмальгаузен, являются внутренние факторы, их нужно
искать внутри самой системы.
Постоянство клеточного цикла у растущих независимо
друг от друга одноклеточных организмов, привело некоторых исследователей к
ошибочному выводу о бессмертии в мире одноклеточных. На самом деле здесь мы
имеем дело с бессмертием, и то относительным, популяций одноклеточных организмов. Когда мы говорим о постоянстве клеточного
цикла у одноклеточных, мы как бы не заглядываем внутрь
цикла и не задаемся вопросом: а как происходит нарастание массы внутри этого
цикла, равномерно или неравномерно. Это мы можем сделать, основываясь на
экспериментальных данных акад. И. И. Шмальгаузена по измерению нарастания массы
3-х одноклеточных организмов: бактерии Micrococcus, дрожжевой клетки и парамеции. Эти данные, выполненные
в 1926 г., до сих пор являются единственными в отечественной и мировой
литературе. Речь идет о том, как нарастает масса отдельной клетки. Анализ этих данных говорит о том, что в течение
клеточного цикла рост происходит по тому же закону, что и рост многоклеточных
организмов, т.е. по закону удвоения массы с линейно возрастающим периодом.
Постоянство же клеточного цикла у популяций одноклеточных сохраняется потому,
что после митоза дочерние клетки омолаживаются и получают тот потенциал роста,
который имела материнская клетка в начале своего роста. Этого омоложения не
происходит в многоклеточном организме; причина этого заложена в том, что
дочерние клетки после митоза не отделяются друг от друга, как в популяции одноклеточных, а остаются связанными в единую массу. В
многоклеточном организме расходуется потенциал, заложенный в зиготе, от
удвоения к удвоению он становится все меньше, и как только израсходуется полностью,
рост прекращается.
По количественному росту растений экспериментальные
данные очень скудные, но и они дают возможность прийти к заключению, что рост
растений происходит по тому
же закону.
Итак, анализ
экспериментальных данных по
количественному росту разных
биологических объектов приводит
нас к выводу,
что рост в живой природе происходит по одному и тому же
простому закону. Выразим это в виде математической формулы.
Мы видели, что зависимость a=f(t) является прямой линией. Из элементарной математики
известно, что уравнение прямой линии в координатах (a,t) имеет вид:
a=a0+kt, где k - угловой коэффициент наклона прямой к горизонтальной
оси, а a0 - начальное значение a при t=0. Подставив
это значение a в формулу (3) вместо
a(t), получим формулу нарастания массы живого организма:
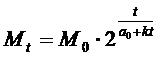 (4)
(4)
Как частный случай этой формулы при k=0 получим
формулу роста популяции независимо растущих друг от друга одноклеточных
организмов.
Еще раз обратим внимание на то, что биологический
смысл параметров a0 и
k нам
не известен. По этой причине формула (4) имеет чисто теоретическое значение.
Исследуя нормальный рост, нельзя не задаться вопросом:
что меняется в формуле (4) в случае злокачественного роста?
Для ответа на этот вопрос, нужно сравнение количественных
данных по злокачественному росту с экспериментальными данными по нормальному
росту. Таких данных в литературе нет. Я благодарю заведующего диагностическим
отделением центра детской онкологии, гематологии и иммунологии в Белоруссии,
кандидата мед. наук И. В. Бегуна за предоставленные им исходные
данные по росту эмбриональных опухолей почек и печени. Анализируя эти данные,
И. В. Бегун вместе со своими
сотрудниками пришел к выводу о том, что указанные опухоли замедляют свой рост с
возрастом, также как нормальные ткани, т.е. в выражении периода a=a0+kt слагаемое kt присутствует как при нормальном, так и при
злокачественном росте. Математический анализ собранных данных подтверждает, что
это действительно так. Более того, коэффициент k практически одинаков как для одного, так и для
другого роста. Получается, что этот параметр не имеет отношения к злокачественному
росту. Тогда напрашивается вывод: злокачественный рост связан с параметром a0, а
именно, с его уменьшением. Этот параметр
не зависит от возраста, а зависит от каких-то иных, пока нам неизвестных
причин. Если в формуле:
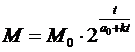 параметр a0 уменьшится, хотя бы незначительно, то число удвоений
параметр a0 уменьшится, хотя бы незначительно, то число удвоений ![]() за
одно и то же время увеличится. Нормальный рост характеризуется постоянством
параметров a0 и k на
всем протяжении роста данного биологического объекта. Представим себе, что
среди нормальных клеток появилась клетка с уменьшенным параметром a0. Клеточный
цикл такой клетки станет короче, потомство такой клетки будет быстрее наращивать
массу по сравнению с соседними,
нормальными клетками. Отсюда все те последствия, которые характеризуют
злокачественный рост: образовавшаяся популяция, обладающая большим ростовым
потенциалом, теснит соседние клетки, питается за их счет. Лишенные питания
соседние клетки отмирают и разлагаются.
за
одно и то же время увеличится. Нормальный рост характеризуется постоянством
параметров a0 и k на
всем протяжении роста данного биологического объекта. Представим себе, что
среди нормальных клеток появилась клетка с уменьшенным параметром a0. Клеточный
цикл такой клетки станет короче, потомство такой клетки будет быстрее наращивать
массу по сравнению с соседними,
нормальными клетками. Отсюда все те последствия, которые характеризуют
злокачественный рост: образовавшаяся популяция, обладающая большим ростовым
потенциалом, теснит соседние клетки, питается за их счет. Лишенные питания
соседние клетки отмирают и разлагаются.
Итак, вывод: злокачественный рост связан с уменьшением
параметра a0 в формуле нарастания массы. Но почему это происходит,
математика ответа не даст. Она только подсказывает, где нужно искать.
Проблема злокачественного роста, да и нормального
роста – проблема междисциплинарная, требующая участия многих наук.
Думаю, что правильная, соответствующая реальности
теория злокачественного роста может быть создана путем объединения результатов,
полученных разными науками. Нужно выдвигать гипотезы, проверять их на соответствие
эксперименту, отбрасывать те, которые ему не соответствуют, выдвигать новые, и
т. д., пока не будет найдена та, которая полностью соответствует наблюдаемым
фактам и эксперименту.
About growth of live organisms
Sedova G. P.
In this article I wish to
sum up the result of the researches on proliferation
live organisms which are more correct for carrying to applied mathematics.
I will concisely state conclusions from the articles devoted to
this subject and I will try to show
that way which brought me to them.
Key words: proliferation live organisms.
Орел.