УДК 612.821
К 70-летию
со дня рождения академика
РАМН, профессора
Константина
Викторовича СУДАКОВА

СУДАКОВ Константин Викторович – родился 6 июля 1932 года. Окончил 1-й Московский медицинский институт им. И. М. Сеченова. Докторскую диссертацию «Нейрофизиологические механизмы пищевого возбуждения» защитил в 1965 году. В течение 1966-1967 гг., будучи стипендиатом Всемирной организации здравоохранения, работал в лаборатории Эволюции лимбической системы Национального института здоровья (Бетезда, США). С 1967 по 1974 гг. – профессор кафедры нормальной физиологии 1 ММИ им. И. М. Сеченова, с 1974 г. и по настоящее время – директор ГУ НИИ нормальной физиологии им. П. К. Анохина РАМН, заведующий кафедрой нормальной физиологии ММА им. И. М. Сеченова. Константин Викторович СУДАКОВ является академиком-секретарем Отделения медико-биологических наук РАМН, сопредседателем Российской секции Международной академии наук, заслуженным деятелем науки РФ.
За выдающийся вклад в развитие современной физиологической науки неоднократно награждался правительственными наградам, памятными медалями, дипломами многих академий, университетов различных стран мира.
Константин Викторович СУДАКОВ в 1982 г. посетил Смоленскую государственную медицинскую академию, читал лекции студентам, выступил с большим докладом перед научной общественностью вузов г. Смоленска. «Актовая речь» Константина Викторовича по случаю 100-летнего юбилея академика П. К. Анохина была опубликована в одном их номеров электронного медико-биологического журнала «Математическая морфология».
В канун 70-летия со дня рождения и 45-летия научно-педагогической деятельности коллективы кафедр физиологии СГМА, СГИФК, редколлегия электронного медико-биологического журнала СГМА «Математическая морфология» сердечно поздравляют Константина Викторовича СУДАКОВА и желают ему доброго здоровья, душевных сил, долгих лет жизни, успехов в плодотворной работе по расшифровке тайн системной организации мозга и окружающего мира.
Заведующий кафедрой физиологии СГМА В. А. ПРАВДИВЦЕВ
Заведующий кафедрой физиологии СГИФК Э.
А. ГОРОДНИЧЕНКО
Профессор кафедры физиологии СГМА С. Б. КОЗЛОВ
Главный редактор электронного
медикобиологического журнала СГМА
«Математическая морфология» В. А. ГЛОТОВ
Выдающемуся
русскому физиологу Константину
викторовичу Судакову – посвящается
НЕЙРОННЫЕ КОРРЕЛЯТЫ АКЦЕПТОРА РЕЗУЛЬТАТОВ ДЕЙСТВИЯ
© 2002 г. В. А.
Правдивцев, С. Б. Козлов, Н. М. Осипов
Кафедра нормальной физиологии
Смоленской государственной медицинской академии
Россия, 214019, Смоленск, ул.
Крупской, 28
В работе
представлены данные, иллюстрирующие
нейрофизиологические особенности
эфферентно-афферентной конвергенции на корковых нейронах.
Установлено, что в процессе сочетаний
раздражения аксонов пирамидного тракта (ПТ) с электрокожным
подкреплением часть изученных нейронов изменяет
параметры вызванной активности на стимуляцию
ПТ, при этом вызванные ПТ-реакции начинают
воспроизводить структуру ответов на подкрепляющие воздействия.
Наиболее динамичные перестройки ПТ-ответов
получены в опытах с включением исследуемых
нейронов в биотехнический комплекс с обратной
связью – «нейрон–ЭВМ–стимулирующие
устройства–животное», фактически
представляющий аналог
натуральной функциональной системы.
Обсуждается
роль эфферентно-афферентной конвергенции на нейронах ЦНС в
развертывании акцептора результатов действия функциональной системы
натурального поведенческого акта.
Ключевые
слова:
пирамидный тракт, эфферентно-афферентная конвергенция, акцептор
результатов действия.
С
позиции теории функциональной системы П. К. Анохина, архитектоника
целенаправленного поведенческого акта
выступает как производное множества
соподчиненных узловых механизмов, начиная с
афферентного синтеза и кончая достижением конечных полезных
результатов и их оценкой [1-4]. В целом, общая
организация циклически замкнутого контура функциональной
системы возражений не вызывает [6, 16], однако
принципы работы составляющих ее
механизмов, особенно центрального
уровня – предмет споров
и дискуссий.
По-прежнему
актуален вопрос о нейрональном генезе акцептора
результатов действия, формирующегося в
функциональной системе в виде своеобразного
контроллера, фиксирующего на своих элементах
модель будущих результатов
поведенческого акта. Предполагается, что
эта модель направляет развитие функциональной
системы путем
сопоставления ее
нейронных производных с
афферентацией о параметрах реально полученных
результатов кванта поведенческого
акта [11, 12].
Наиболее
известной гипотезой, постулирующей вовлечение нейронов в акцептор
результатов действия, является гипотеза,
выдвинутая П. К.
Анохиным [2-4] и
дополненная К. В.
Судаковым [11-14]. Согласно этой гипотезе,
ключевым моментом многоэтапного формирования
акцептора результатов действия является
распространение по корково-подкорковым
структурам ЦНС эфферентных возбуждений
корковых мотонейронов – нейронов пирамидного
тракта (ПТ) [18, 21-24, 26]. Субстрат
распространения - аксонные коллатерали.
Учитывая, что коллатеральные эфферентные
возбуждения копируют
моторную программу, извлекаемую из памяти,
делается вывод о том, что в дальнейшем
коллатеральные копии моторных возбуждений
считываются и запоминаются в нейрональном
комплексе акцептора результатов действия.
Допускается, что активированный коллатеральными
возбуждениями акцептор результатов действия,
сохраняя в закодированном виде копию моторной
программы, фактически
модель будущих результатов, сопоставляет
и сравнивает ее с обратной афферентацией. Таким
образом, принимается очередное предположение о
том, что морфо-функциональным модулем акцептора
результатов действия является ансамбль
вставочных нейронов, а его элементарным
операционным процессом выступает
эфферентно-афферентная конвергенция,
обеспечивающая интеграцию возбуждений
различной системной направленности и
информационной значимости [4,
11, 12].
Настоящее
исследование предпринято с целью проверки
некоторых положений изложенной гипотезы. В
частности, нас
интересовали формы нейрофизиологических
проявлений эфферентно-афферентного
взаимодействия возбуждений, воспроизводящего
на нейронах соотношения, характерные для
системной активности акцептора результатов
действия. Конечная цель экспериментов
заключалась в изучении динамических компонентов
вызванных коллатеральных ответов нейронов
теменной ассоциативной коры на стереотипную
стимуляцию ПТ, подкрепляемую соматическими
электрокожными раздражениями. Включение
подкрепляющего раздражения
причинно увязывалось с качеством ответов
нейронов на поступление к ним коллатеральных
эфферентных возбуждений ПТ, что позволяло
имитировать определенные закономерности,
присущие избегательным поведенческим актам.
Последнее обеспечивалось
введением регистрируемого нейрона в гибридный
биотехнический комплекс [8, 15] в составе
«нейрон-ЭВМ-стимулирующие
устройства-подопытное животное».
МЕТОДИКА
Опыты проводили на кошкаx, массой 3,5-4 кг. В ходе
подготовительного этапа в условиях
этаминал-натриевого наркоза (30-35 мг/кг) животным
рассекали мягкие ткани, в черепе просверливали
трепанационные отверстия. Края ран
инфильтрировали 0,5% р-ром новокаина. Животных
интубировали, обездвиживали
миорелаксантами и переводили на управляемое
дыхание.
Для
введения нихромовых электродов в пирамиды
продолговатого мозга использовали вентральный
доступ через затылочно-основную кость.
Стимуляцию аксонов ПТ осуществляли
прямоугольными импульсами тока амплитудой 5-7
В (0,05мс), частотой – 200/с. Считали, что электроды
находятся в ПТ, если одиночный электрический
импульс вызывал в передних областях коры
фокальный антидромный потенциал. Электрокожные
воздействия наносили через игольчатые
электроды, введенные в подушечки
контралательной задней лапы (20-40 В, 0,5 мс)
животного.
Внеклеточное
отведение потенциалов одиночных нейронов
осуществляли стеклянными микроэлектродами,
заполненными 2 М р-ром NaCl.
Сопротивление электродов по постоянному току
не превышало 10 МОм. Черепное отверстие в зоне
отведения нейронов после установки
микроэлектрода заливали 3% р-ром агара,
приготовленным на физиологическом растворе.
Регистрацию нейронов начинали через 6-8 ч после
введения этаминал-натрия. Система усиления и
отведения внеклеточных потенциалов нейронов
была стандартной.
Анализ
активности нейронов по показателям растровых
нейронограмм и усредненных перистимульных
гистограмм проводили с помощью лабораторной ЭВМ.
Специализированные программы обработки
нейронной активности, устройства сопряжения
усилителей биопотенциалов, стимуляторов с ЭВМ разработаны в СКБ СФ МЭИ.
Опыт сводился к следующему.
После предварительной оценки фоновой и
вызванной активности нейрона на
вышеуказанные воздействия, а также на
вспышки света, звуковые щелчки, вестибулярные
воздействия, опыт переводился в автоматический
режим. При этом ЭВМ, запуская
устройства регистрации и стимуляции, проводила подсчет числа
импульсов на протяжении времени раздражения (500
мс) ПТ. Полученная величина сравнивалась с
эталоном первичных данных о нейронной
активности. В случае превышения нейроном
эталонного уровня активности во время
стимуляции ПТ, ЭВМ блокировала включение
электрокожного подкрепления.
Результаты исследований
Всего в
теменной ассоциативной коре (5 поле по Бродману)
было зарегистрировано 164 нейрона. Из них 68
реагировали на ритмическую стимуляцию
пирамидного тракта (ПТ). Латентные периоды
реакций у разных нейронов варьировали от 10 до 200
мс.
Из
группы нейронов, реагировавших на стимуляцию ПТ,
56 (82%) изменяли свою активность на электрокожное
раздражение, 39 нейронов (57%) отвечали на звуковые
щелчки или вспышки света, 31 нейрон (46%)
реагировал на вестибулярные воздействия при
угловых перемещениях животного во фронтальной
плоскости.
В
представляемой серии опытов подробно
анализируется активность 37 нейронов, отобранных
нами преимущественно по критериям фазной
реактивности на электрокожное раздражение, а
также реактивности на ритмическую стимуляцию
аксонов ПТ.
Перед
проведением программы основной части опытов во
всех случаях проводилось
тестирование ответов нейронов на
последовательные изолированные стимуляции
аксонов ПТ (не менее 20) с интервалами
30-45 с без подкрепляющих воздействий. Типичной реакцией при
выполнении такого тестирования было частичное
угашение ответов. В 5-ти случаях
угашение было
полным. После стабилизации ответов на стимуляцию
ПТ приступали к обусловливанию путем
сочетаний стимуляции ПТ с оставленным
электрокожным подкреплением.
На рис. 1
представлена динамика нейронной активности в
опыте при проведении обусловливания. Исходный ответ нейрона на
раздражение ПТ (а-) невыразителен, отмечается незначительная активация. В процессе
проведения сочетаний (b-) в
четвертом предъявлении раздражителей отмечаем
стабилизацию ответа на стимуляцию ПТ, появление в его составе
ранних компонентов. На данном этапе
обусловливания продолжение опыта проходило в
режиме ЭВМ-управления. Предполагалось, что
электрокожное раздражение будет устранено,
если за 500 мс раздражения ПТ ЭВМ зафиксирует
генерацию нейроном 25 или
более разрядов.
Результаты
очередного фрагмента опыта –
с- и
d-.
Из них на с- отдельно
выделены положительные реализации (всего 11), в
которых нейронная импульсация во время
стимуляции ПТ превысила установленный порог,
тогда как на d- собраны
отрицательные реализации (всего 9), в которых
порог вызванной активности превышен не был,
поэтому они сопровождались электрокожным
подкреплением. Выявленные различия вызванной
нейронной активности на нейронограммах с- и d- очевидны.
В
дальнейшем констатируем,
что количество положительных реализаций при
продолжении опыта существенно повышается. Так, в
блоке нейронограмм е- из 20 предъявлений
раздражителей уже в 15 случаях отмечается
«избегание» электрокожного подкрепления.
Усредненный ответ на стимуляцию ПТ в целом
усилен, трансформирован,
причем все это происходит при уменьшении числа
подкрепляющих воздействий. Отличительной
особенностью обусловливания в опытах с
избегательным поведением, это отмечают многие,
является трудность угашения образованных
навыков. Аналогичный феномен наблюдался нами
практически во всех ЭВМ-управляемых опытах, т. к.
оказалось, что период угашения приобретенных
вызванных ответов нейронов обычно увеличивался
в 1,5-2 раза в сравнении с экспериментами, в которых
электрокожное подкрепление вводилось вне
зависимости от качества упреждающей реакции
нейрона.
На
фрагменте f- рис. 1 приводятся растровые нейронограммы, отражающие
активность нейрона на заключительном этапе
угашения после проведения основной части опыта.
Лишь после 35 изолированных стимуляций ПТ
выраженность ответов оказывается сопоставимой с
исходными ответами.
Рассмотрим
детальнее содержание тех опытов, в которых
активность нейронов
регистрировалась на протяжении длительного
времени при многократных изменениях условий
запуска электрокожного подкрепления. По нашему
мнению, в таких опытах
можно ориентировочно оценить динамизм нейронной
активности в процессе преодоления
рассогласования при непрогнозируемом
поступлении к нейрону электрокожных
подкрепляющих возбуждений.
На рис. 2-а - исходная активность
нейрона. Ответ на стимуляцию ПТ слабо выражен. На
рис. 2-b - тот же нейрон в
процессе сочетаний ПТ с электрокожным
подкреплением. На растровых нейронограммах с седьмого сочетания
отмечаются выраженные изменения активности во
время стимуляции ПТ. Усредненный ответ всего
опытного блока в сравнении с начальным
достоверно усилен. Констатируем значительный рост фоновой активности
нейрона – с 1 имп/с до 21,1 имп/с.
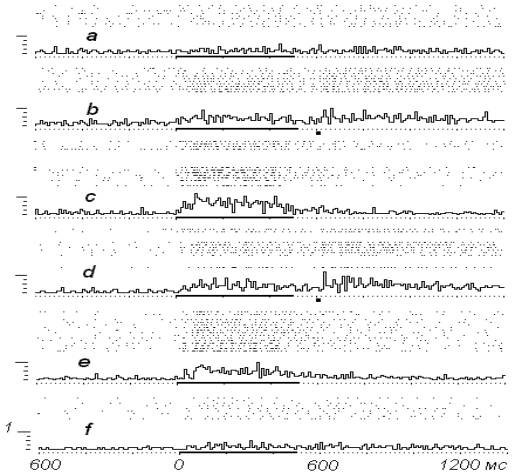
Рис. 1.
Реакции нейрона на стимуляцию ПТ в условиях
ЭВМ-управляемого эксперимента.
a-
исходный ответ на стимуляцию ПТ, b- ответы при сочетании
стимуляции ПТ и электрокожного подкрепления, c-, d- то же в условиях
ЭВМ-управляемого эксперимента (в блоке с- из 20 предъявлений раздражителя
изолированно представлены положительные
реализации, на d- отрицательные реализации,
сопровождающиеся электрокожным подкреплением), e- то же при продолжении эксперимента (представлены
положительные реализации), f- ответы
нейрона после угашения. По вертикали - число
импульсов в бине (10 мс), по горизонтали - время, мс.
Горизонтальной
чертой отмечен период раздражения
ПТ, точка - электрокожное подкрепление.
На рис. 2-с представлена активность
того же нейрона при переходе в режим работы в
составе биотехнического комплекса. Начальный
порог выключения электрокожного подкрепления
установлен на уровне 17 разрядов за 500 мс
раздражения ПТ.
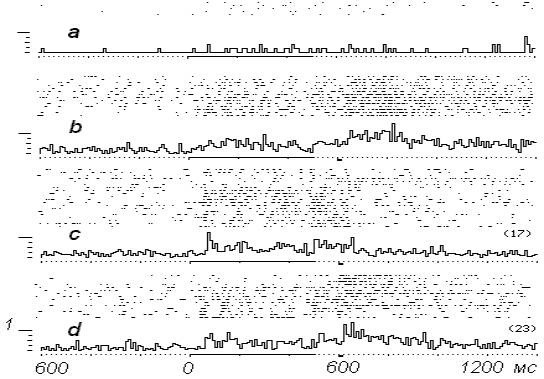
Рис. 2.
Динамика нейронной
активности в ЭВМ-управляемом эксперименте с
варьированием порога выключения электрокожного
подкрепления. а- исходный ответ, b- в ходе сочетаний стимуляции ПТ с электрокожным
подкреплением, c-
управление в опыте передано ЭВМ,
порог выключения подкрепления
- 17 разрядов /500 мс, d- то же при повышении
порога до 23 разрядов/500 мс.
Итак, в
первых двух предъявлениях раздражителей порог
был преодолен, что
привело, очевидно, из-за отсутствия подкрепления,
к снижению вызванной активности. Порог
преодолеваться перестал, и последующие четыре
стимуляции ПТ прошли в сопровождении
электрокожного подкрепления. В результате
сочетаний стимуляции ПТ с электрокожным
подкреплением должный уровень вызванной
активности восстановился, так что порог выключения подкрепления теперь уже
преодолевается во всех пробах до конца опытного
блока (-с).
Отметим
изменение структуры ответа нейрона на
стимуляцию ПТ – исходный тонический ответ
преобразовался в фазный. В его составе появился коротколатентный
компонент, хорошо выявляемый при избирательном
усреднении положительных реализаций
настоящего фрагмента опыта. Подчеркнем, что
преобразование вызванного ответа нейрона
происходило параллельно с изменениями фоновой
активности. Последняя снизилась
до 18,1 имп/с.
На рис. 2-d наблюдаем новые сдвиги
вызванной активности нейрона при очередном
изменении опытной ситуации, когда порог
выключения электрокожного подкрепления был
повышен с 17 до 23 разрядов. Фиксируем, что в первых
четырех воздействиях на ПТ порог не
преодолевается, тогда как, начиная с пятого, из-за
повышения реактивности нейрона, порог стал преодолеваться почти всех
предъявлениях раздражителей (9 из 11) до конца
опытного блока. Усредненный ответ данных
реализаций представлен на рис.
3-а, -b.
Рис. 3 (c-, d-, e-) демонстрирует
развитие экспериментальной ситуации при повышении
порога с 23 до 25 разрядов за 500 мс стимуляции
ПТ.
В
воздействиях на ПТ с первого по четвертое порог
не преодолевается, животное всякий раз получает
электрокожное подкрепление. Обращает на себя
внимание тот факт, что невозможность преодоления
в первых реализациях установленного порога,
вызывает скачкообразное изменение типа фоновой
активности нейрона, в ее составе начинает
доминировать пачечный ритм. Частота фоновой
активности значительно
возрастает, маскируя ответы на стимуляцию ПТ
(рис. 3-с).
Как выяснилось, усиленный фоновый ритм
нейрона обеспечил требуемое число разрядов во
время раздражения ПТ для устранения
подкрепления в предъявлениях с пятого по
восьмое. Последнее обстоятельство немедленно
отразилось на фоновой активности. Она снизилась
до 9,4 имп/с и в дальнейшем преодоление
порога, устраняющее электрокожное подкрепление,
обеспечивалось исключительно за счет разрядов
вызванных ПТ-ответов (рис.
3-d).
На примере рассмотренного
опыта можно сделать предварительные заключения
о различных механизмах преодоления нейронами
состояний рассогласования при отрицательном
соотношении наличной вызванной активности на
стимуляцию ПТ и эталона (порога) выключения
электрокожного подкрепления.
Первый
механизм обнаруживает себя преимущественно в
перестройках частоты фазных ответов на
раздражение ПТ до уровня, достаточного для
преодоления порогов выключения подкрепления.
Данный механизм мы наблюдали во всех опытах,
обычно на начальных этапах ЭВМ-управляемых
опытов.
Второй механизм проявляется в
изменениях фоновой активности нейронов. При этом
часто появляется пачечная ритмика, частотные
характеристики которой, перекрывая частотные
характеристики вызванных ПТ-ответов,
обеспечивают положительный эффект выключения
подкрепления. Реализацию данного механизма мы
наблюдали в 9-ти из 26-ти случаев, обычно на поздних
стадиях развития опытной ситуации, когда
установленные пороги значительно превышали
показатели вызванных ПТ-ответов.
Третий механизм – смешанный,
отражающий перестройки и вызванных ответов, и
фоновой активности нейрона на самых различных
этапах проведения опытов, как
это показано на рис. 4, 5 (16 случаев из 26).
На рис. 4-а видно, что нейрон
исходно реагировал на раздражение ПТ
невыразительно, окончание стимуляции
сопровождается угнетением активности.
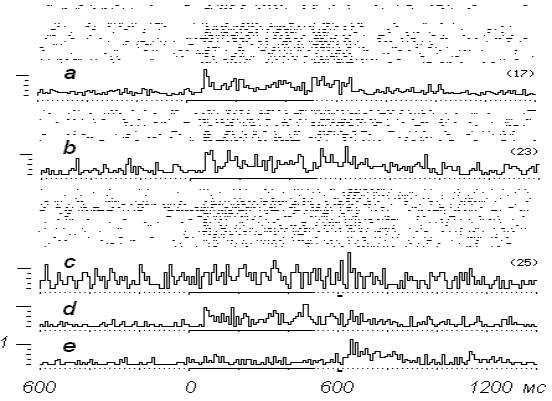
Рис. 3. Динамика
нейронной активности в ЭВМ-управляемом эксперименте с
варьированием порога
выключения электрокожного подкрепления (продолжение опыта,
представленного на рис 2). а-,
b- изолированное представление положительных реализаций из блоков
нейронограмм рис. 3-с, d-. Далее - на растровых
нейронограммах - активность нейрона в блоке 20 предъявлений
раздражителей в условиях повышения порога выключения электрокожного подкрепления до 25 имп/500 мс, с- перистимульная усредненная активность нейрона в предъявлениях
раздражителей (по
растровой нейронограмме) с 5 по 8 включительно, d- усредненная гистограмма
положительных реализаций (предъявления 9-20),
представленных на растровых
нейронограммах, e- то же для отрицательных реализаций (первые 4 предъявления).
В
результате сравнительно большого числа
сочетаний (20) раздражения ПТ с электрокожным
подкреплением не было обнаружено какой-либо
направленной динамики (рис. 4-b).
Положение изменилось
при переходе к режиму ЭВМ-управляемого этапа
эксперимента.
В блоке
растровых нейронограмм рис. 4-с отмечаем, что в 11
случаях установленный порог - 15 разрядов за
период раздражения ПТ (500 мс) - был преодолен.
Усредненные положительные
и отрицательные реализаций показаны на
гистограммах d- и e-.
По ходу
опыта отмечаем наращивание
количественных показателей ответа нейрона,
отчетливое изменение его паттерна, в составе
которого появляются новые фазы активаций - f-, g-.
В
последующем (рис. 5, а-, b-) при
очередном повышении порога «избегания»
электрокожного подкрепления до 20 разрядов вновь
отмечаем дальнейшие
перестройки активности, причем проявляются они
не только в изменениях вызванной, но и фоновой
активности. Обращает на себя внимание
сформировавшийся в составе вызванного ответа
комплекс, фактически воспроизводящий ответ на
электрокожный раздражитель после окончания
стимуляции ПТ.
При
новом изменении порога выключения
электрокожного подкрепления (23 разряда)
наблюдаем усиление, в основном,
предстимуляционной активности
нейрона (рис. 5-d). В целом из блока 20 предъявлений
раздражителей в 15 порог
выключения электрокожного подкрепления был
преодолен.
Из исследованных 37 нейронов, 26
продемонстрировали изменения активности,
которые можно было трактовать как проявление
обучения, направленного на минимизацию
биологически отрицательного электрокожного
подкрепления в составе функционального
биотехнического комплекса. Форма сдвигов
нейронной активности оказалась различной, при этом типичным
вариантом был переход тонических ответов на
стимуляцию ПТ к фазным, сопоставимых по
выраженности и конфигурации с реакциями
нейронов на электрокожные раздражения. Из числа
нейронов, не обнаруживших сдвигов в
ЭВМ-управляемых опытах (11), выделялись нейроны (6)
с правильной, регулярной ритмикой и
маловыразительными тоническими ответами на
стимуляцию ПТ.
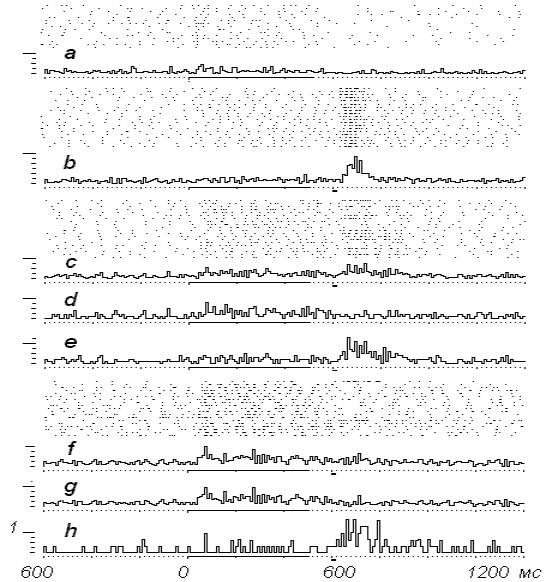
Рис. 4. Динамика нейронной
активности в ЭВМ-управляемом эксперименте. а-
на растровых нейронограммах и
усредненной гистограмме - реакция нейрона на
раздражение ПТ, b- в
процессе сочетаний стимуляции
ПТ с электрокожным
подкреплением, с- нейрон в
ЭВМ-управляемом режиме опыта,
порог 15, d- усредненные
положительные реализации (11), е-
усредненные отрицательные реализации, f-
продолжение опыта, g-
усредненные положительные реализации (17),
h- отрицательные реализации.
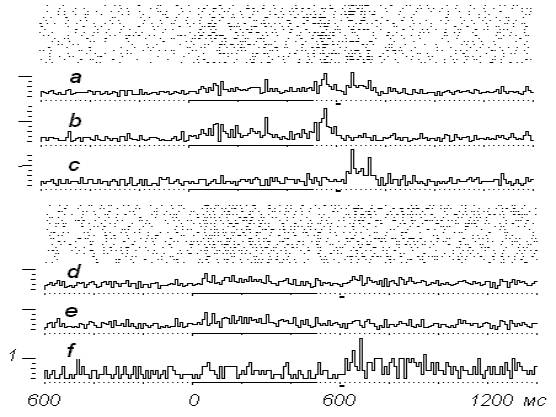
Рис.5.
Продолжение опыта, представленного
на рис. 4. а- порог выключения
электрокожного подкрепления повышен до 20/500 мс, b- усредненная гистограмма положительных
реализаций (11), с- усредненная
гистограмма отрицательных реализаций, d- продолжение опыта, е- положительные реализации (15), f-
отрицательные реализации (5).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализируя
данные проведенной серии опытов, констатируем,
что в целом они не противоречат гипотетическим
представлениям об акцепторе результатов
действия, основанным на понимании особой
значимости эфферентно-афферентной конвергенции в ЦНС.
Что
обращает на себя внимание в первую очередь?
Отметим
синаптическую
активацию значительной
части нейронов
теменной ассоциативной коры под влиянием
стимуляции аксонов,
образующих ПТ.
Подчеркнем, что большинство исследованных нейронов,
реагировавших на стимуляцию
эфферентных проводников ПТ,
отвечали и на
афферентные возбуждения различной
сенсорной
модальности. В
дальнейшем были выявлены факты,
указывающие на наличие более
существенной связи
между коллатеральными эфферентными
и афферентными возбуждениями. Речь
идет о фактах, свидетельствующих о том, что
нейроны, перерабатывая коллатеральные
эфферентные возбуждения, одновременно
реализуют процесс оценки сигнальных
составляющих в структуре этих возбуждений. Об
этом говорит феномен пластичности нейронных
ответов при последовательной изолированной
стимуляции ПТ с неизменными параметрами.
Пластичность в форме ослабления или полного угашения
вызванных ответов интерпретировалась нами как проявление
неассоциативного обучения [9, 10]. С позиций
возможной причастности нейронных объединений
теменной коры к акцептору результатов действия
кажется допустимым, что отсутствие в составе
коллатеральных эфферентных возбуждений
сигнальных составляющих приводит к
постепенному исчезновению вызванных ответов
этих объединений.
Иную
картину наблюдали в ситуации обусловленного
взаимодействия эфферентных коллатеральных
возбуждений с афферентными подкрепляющими
возбуждениями.
В работах
К. В. Судакова, посвященных анализу
принципиальной схемы формирования акцептора
результатов действия, роль фактора, фиксирующего
специфический активированный статус
пространственно-временного ансамбля нейронов
акцептора результатов действия,
отводится подкрепляющим возбуждениям [12, 13]. Подчеркивается, что при
повторении элементарного цикла системной
деятельности соответствующий нейрональный
статус акцептора
результатов действия воспроизводится –
первично за счет процессов афферентного
синтеза, окончательно – в результате введения в
акцептор результатов действия сигнальной
информации, представленной коллатеральными
эфферентными возбуждениями.
Какие
опыты позволяют оценить роль подкрепляющих
возбуждений применительно к обсуждаемой
динамике эфферентно-афферентных процессов
в акцепторе результатов действия?
Прежде
всего, это опыты, в
которых было установлено, что
направленные перестройки фазных
нейронных ПТ-ответов,
воспроизводящие паттерн, характерный для
ответов этих же нейронов на подкрепляющие
воздействия, формируются при сочетаниях
стимуляции аксонов ПТ с подкрепляющими
возбуждениями, т. е. при обучении. Наиболее
очевидные примеры перестроек ПТ-ответов
получены нами в опытах с включением исследуемых
нейронов в биотехнический
комплекс, представляющий собой фактически
аналог натуральной функциональной системы. При этом констатировали,
что как только вероятность биологически
значимого афферентного воздействия на животное
жестко ставилась в зависимость от маркированных
показателей фазных ПТ-ответов, в большинстве
случаев происходили быстрые и прогнозируемые
изменения нейронной ритмики, особенно в ответах
на стимуляцию ПТ. Оказалось, что многократное
варьирование условий введения подкрепления
всегда приводило к многократным трансформациям
и спонтанной, и вызванной активности нейронов на стимуляцию ПТ.
Вполне
уместным представляется вопрос о том, на какой
основе реализуется фазное изменение
интервально-частотных характеристик ответных
реакций нейронов при поступлении к ним
коллатеральных эфферентных возбуждений в
процессе их взаимодействия
с афферентными подкрепляющими
возбуждениями? Иначе, как организован
принципиальный механизм опережающего
воспроизведения паттерна подкрепляющих –
результативных возбуждений в акцепторе
результатов действия во время считывания
нейронами коллатеральных эфферентных
возбуждений?
Исходя из
того, что качество ответов на стимуляцию ПТ –
производное обучения, допускаем, что
воспроизведение паттерна
подкрепляющих возбуждений в акцепторе
результатов действия под влиянием
коллатеральных возбуждений ПТ является
следствием актуализации на нейронах-мишенях
вновь образованной энграммы,
оперативно преобразующей активность
генераторного пункта этих нейронов [7]. С
учетом данного предположения, тип влияний
коллатеральных возбуждений
корковых мотонейронов на нейрональный аппарат
акцептора результатов действия применительно к
адаптивному режиму активности функциональной
системы может быть определен как пусковой.
Вполне возможно, что инициация пуска энграммы
происходит в ответ на поступление к нейронам
строго фиксированных по интервальным параметрам
совокупных потоков коллатеральных
эфферентных возбуждений.
К
настоящему моменту проведено большое количество
исследований, в которых активность
структур разного уровня ЦНС изучалась
в ходе натуральных
поведенческих актов [5, 17, 19, 20, 25]. Результаты
таких экспериментов, с одной стороны,
предоставили в распоряжение аналитиков
разнообразный фактический материал, с другой –
продемонстрировали сложность сведения
материала в единую структурно-логическую
конструкцию. По этой причине сохраняют
актуальность простые эксперименты, в которых
изучение функциональной иерархии связей
нервных структур, механизмов, потенциально
участвующих в обеспечении поведенческого акта,
проводится в условиях, уменьшающих
неопределенность в интерпретации данных.
Именно с этих позиций мы рассматриваем место
нашей работы в ряду работ, посвященных
закономерностям центрального обеспечения поведенческих актов, в
частности, закономерностям
построения акцептора результатов действия.
СПИСОК
ЛИТЕРАТУРЫ
[1] Анохин П. К. Узловые вопросы
в изучении высшей нервной деятельности //
Проблемы высшей нервной деятельности. Изд-во АМН
СССР, М.: 9–128. 1949.
[2] Анохин П. К. Кибернетика и
интегративная деятельность мозга //
Кибернетические аспекты интегральной
деятельности мозга. 18 Международный
психологический конгресс. Наука,
М.: 3–20. 1966.
[3] Анохин П. К. Биология и
нейрофизиология условного рефлекса. Медицина, М. 1968.
[4] Анохин П. К.
Методологическое значение кибернетических
закономерностей // Очерки по физиологии
функциональных систем. Медицина, М.: 322–346. 1975.
[5] Батуев А. С.
Принцип доминанты в организации
целенаправленного поведения // Физиол. журн. им.
И. М. Сеченова. 80 (6) : 94–100. 1994.
[6] Вартанян Г. А., Пирогов
А. А. Нейробиологические
основы высшей нервной деятельности. Наука, Л. 1991.
[7] Пирогов А. А. Нейродинамика
смены энграмм. Наука, С. Пб. 1991.
[8] Полянцев В. А. Постоянный
поиск оптимального режима как принцип
интегративной деятельности мозга
// Кибернетические аспекты интегральной
деятельности мозга. 18 Международный
психологический конгресс, Наука, М.: 166–168. 1966.
[9] Соколов Е. Н. Нейронные
механизмы памяти и обучения.
Наука, М. 1981.
[10] Сторожук В. М. Нейронные
механизмы обучения. Наукова думка, Киев. 1986.
[11] Судаков К. В. Общая теория
функциональных систем. Медицина,
М. 1984.
[12] Судаков К. В. Нейрональные
механизмы системного "кванта" поведения // Нейроны в поведении.
Системные аспекты. Наука,
М: 58–72. 1986.
[13] Судаков К. В. Системный
процесс подкрепления // Журн. высш. нервн.
деятельности. 46 (4) : 643–655.
1996.
[14] Судаков К. В. Рефлекс и функциолнальная
система. Изд-во
Новгородского госуниверситета, Новгород. 1997.
[15] Трубачев В. В. Оперантное
обусловливание импульсной и медленноволновой
активности мозга как операционный принцип
системной интеграции в процессах обучения //
Механизмы интегративной деятельности мозга.
Наука, М. : 227–237. 1981
[16] Шапков Ю. Т., Анисимова Н. П.,
Герасименко Ю. П., Романов С. П. Регуляция
следящих движений. Наука, Л. 1988.
[17] Baker S. N., Spinks R., Jackson A., Lemon R. N.
Synchronization in monkey motor cortex during a precision grip task // J. Neurophysiol. 85
(2) : 869–885. 2001.
[18] Endo K., Araki T., Yagi N. The distribution and pattern of axon branching of pyramidal tract cells // Brain Res. 57(2) : 484-491. 1973.
[19] Evarts E. V., Bizzi R., Burce R. E., DeLong M., Thach W. T. Central control of movement // Neurosci. Res. Program. Bull. 9 : 129–135. 1977.
[20] Evarts E. V., Wise S. P. The cerebral control of
movement // Electroencephalogr. and Clin. Neurophysiol.
61(3): 6-14. 1985.
[21] Giuffrida R., Li Volsi G., Panto M. R., Perciavalle V., Urbano A. Pyramidal influences on ventral thalamic nuclei in the cat // Brain Res. 279 (1-2): 254-257. 1983.
[22] Giuffrida R., Li Volsi G., Maugery G., Perciavalle V. Influences of pyramidal tract on the
subthalamic nucleus in the cat// Neurosci Lett. 54(2-3): 231-235. 1985.
[23] Giuffrida R., Li Volsi G., Maugery G., Perciavalle V. Pyramidal input to basal ganglia in the cat
// Exp. Brain Res. 1986. 61 (3): 645–648. 1986.
[24] Marino J., Canedo A., Aguilar J.// Sensorimotor
cortical influences on cuneate nucleus rhythmic activity in the anesthetized cat//
Neuroscience. 95(3): 657–673. 2000.
[25] Nicolas G., Marchand-Pauvert V., Burke D.,
Pierrot-Deseilligny E.//
Corticospinal excitation of presumed cervical propriospinal neurones and its reversal to
inhibition in humans// J Physiol. 533(3): 903-919. 2001.
[26] Tsumoto T., Nakamura
S., Iwama K. Pyramidal tract control over cutaneus and
kinesthetic sensory transmission in the cat thalamus // Exp. Brain Res. 22(3): 281–294.
1975.
NEURON CORRELATIVES OF THE ACCEPTOR
V. A. Pravdivtsev, S. B. Kozlov, N. M. Osipov
Smolensk State Medical Academy,
Department of Normal Physiology, 214019, Russia, Smolensk, 28, Krupskaya Street
This study presented data illustrating neurophysiological features of the efferent-afferent convergence on the cortex neurons. It was established that during the process of combinations of stimulation of the axons of the pyramidal tract (PT) with electrical skin support, the significant part of studied neurons changed parameters of the evoked activity during PT-stimulation. The most prominent reconstruction of the PT-responses were obtained in experiments with inclusion of study neurons into biotechnical complex with feed back – «neuron–computer–stimulating devices–animal», representing itself an analogue of the functional system.
The role of afferent-efferent
convergence on CNS neurons is discussed in the dynamic deployment of the acceptor of
action results at the functional system of natural behavioral act.
Key words: pyramidal tract,
efferent-afferent convergence, acceptor of action results.