Математическая
морфология.
Электронный
математический и медико-биологический журнал. - Т. 17. -
Вып. 2. -
2018. - URL:
http://www.sci.rostelecom67.ru/user/sgma/MMORPH/TITL.HTM
http://www.sci.rostelecom67.ru/user/sgma/MMORPH/N-58-html/TITL-58.htm
http://www.sci.rostelecom67.ru/user/sgma/MMORPH/N-58-html/cont.htm
CHIRAL PHYSICS OF THE
HUMAN BRAIN
© 2018 Alexander
Kholmanskiya , Nataliya
Zaytsevab
Activation of chiral biomolecules by an external physical factor was
explained by an increase in the effectiveness of melatonin biosynthesis in the
epiphysis of a sleeping person at 2-3 o'clock in the morning. The rate of
propagation of biogenic energy quanta over the lithosphere was estimated and
their solar nature was suggested. The phenomenon of electromagnetic induction,
the magnetic properties of sodium ions and the helicity of the myelin sheaths
of nerve fibers were used to substantiate the inductive mechanism of saltatory
conduction. The equivalent circuit of the neuron membrane was simulated by a
sequence of oscillatory circuits, which allowed to lead the chirality factor
into the algorithm of the operation of neural circuits. They modeled the
structure and properties of an elementary quantum of energy possessing
chirality and proposed rules for their assembly into particles and interactions
with spins and magnetic moments of particles and chiral molecules. Assumed the
participation of solar neutrino energy in the genesis of the morphofunctional
dissymmetry of the human brain.
Key words: chirality;
inductance; energyform; neuron; Ranvier;
vibrational contour.
The ideal prototype of an
artificial intellectual system was and will be the human brain with its unique
ability to heuristically think. The neurophysiology of this ability is based on
the anatomical and functional features of the human brain. These include the
organization and biochemistry of neural networks, as well as physical
mechanisms for the formation, recognition and memorization of new patterns. The
emergence and development of structural and functional features responsible for
the mechanism of thinking can be associated with a mutation of the primacy
genome and subsequent adaptation of its physiology to the action of the
universal biogenic factor (UBF) at a certain stage of evolution. The main
consequence of this mutation was the genesis of anatomy and physiology of the
speech function. Speech acoustics, as an internal factor and UBF, as external,
provided ontogeny and differentiation of a person's intellectual abilities in
accordance with his phenotype [1]. At the same time, visual, olfactory and
tactile sensory play their auxiliary role. The physical nature of UBF has not
been established so far, however, it somehow affected the features of the
mechanisms for processing and synthesizing new information in the human brain.
Obviously, for their understanding, anatomical and biochemical studies of the
brain must be accompanied by a study of the mechanisms of interaction of brain
biosystems with external and internal physical factors - electromagnetic (EM)
and, possibly, neutrino nature: The fundamental dynamic idea of matter ... is so intertwined
with our forms of thinking [2]; the principles of the brain are completely
unusual, ... the dimensionality of his operations is beyond the scope of our
ideas. ... the main thing in the activity of the brain is not a regulatory but
constructive function [3]; The process of structuring ... is a spontaneous
generation of an ordered sequence, a hierarchy of dynamic structures, or
«energyforms» [4].
So far, the functional
asymmetry of the brain, the chirality (optical activity) of biomolecules and
fluid media of the brain, the cooperative effects in homogeneous biosystems,
the phenomenon of EM induction and the impact on the brain of the sleeping
person of geocosmic factors are not properly taken into account when studying
and modeling the mechanism of thinking. Such factors include UBF, and the
genesis of dissymmetry of living systems seems to be associated with it [5]. In
modeling the physics of chiral systems and objects, energy quanta or particles
possessing intrinsic chirality play an important role [6]. For their
construction it is reasonable to use the simplest model of the dynamic form of
matter - energy form (EF) [7]. Despite the fact that there are as
yet no technical methods for direct recording of EF inside and outside the
brain, their effect is manifested in the «vision» of the brain by dreams and
pressure phosphophenes or magnetophosphenes [8]. EF are also involved in the
generation of EM pulses detected by EEG and EMG methods [9].
2.
Modeling of chiral energy quanta
The model of the simplest EF
is constructed taking Maxwell's equations into account for the EM field (1) and
extrapolations of the EM-induction phenomenon (Fig. 1):
rotE
= - дB/дt и rotH = j + дD/дt.
(1)
In (1), E and B are mutually
orthogonal vectors of the strength of the vortex electric and magnetic fields, D = εoεE , B = μoμH,
j is the bias current, and the electrodynamic vacuum constant (εoμo)
and the refractive index of the medium are related to the propagation velocities
of the EM quanta in vacuum (C) and medium (V) by the relationships:
C = (εoμo)–1/2 и V = С(εμ)–1/2 = С/n.
(2)
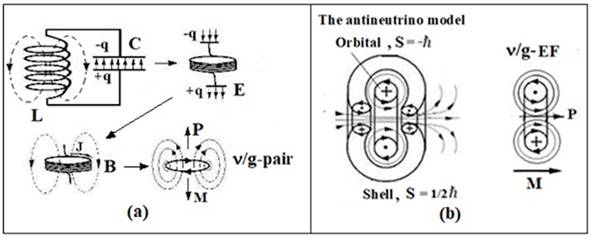
Fig. 1. (a) Vibrational
contour, its transformed forms and their extrapolation to the level of the
energy form (ν/g-pair) having the impulse P,
момент импульса М and the equivalent mass mg. (b)
Antineutrino models (spin S = -ћ/2) and right-handed v/g-EF (P - momentum, M -
angular momentum).
In Fig. 1 shows ν / g-pair simulating a left-handed photon or EM-vortex whose momentum and
angular momentum are directed in opposite directions. From v/g-pair it is
possible to construct vortex models of elementary particles and nuclei [7]. For
example, the antineutrino model will include an ν-shell and a g-orbital with spins ½ and -1, respectively (Fig. 1).
The assembly of ν/g-pair particles or EM energy quanta corresponding to the excited state of
a molecule or a biosystem was represented by the condensation equation N of the
ν/g-pair in the form [8]:
![]() .
(3)
.
(3)
In (3) Rν refers to the characteristic size of a v-vortex of a geocosmic scale, and
re refers to the metric characteristics of an electron or electron orbitals, an
atom, a molecule, or a molecular cluster. In this case, N can reach the
Avogadro number.
The interaction of chiral EFs
with a biosystem consisting of chiral molecules is realized analogously to the
interaction of quasiparticles of magnons with nuclei and electrons having
nonzero magnetic moments. Absorption by a particle or a system of EF particles
will lead to a reorientation of the corresponding magnetic moment and will affect
the spin-spin and spin-orbit interactions of chiral chromophores determining
the anisotropy level of the electronic structure of the entire molecule and its
chirality [7]. This will change the level of cooperation of chiral elements in
homogeneous systems of molecular complexes and supramolecular ensembles in
brain tissues. As a result, the chemical potential will increase, and the
kinetics of biochemical reactions will become more dependent on the chirality
of the molecules5.
3. Chiral physics of the epiphysis.
In the process of
phylogenesis, an anatomical and physiological complex was formed in the human
brain, regulating homeostasis in accordance with the circadian rhythm of
day-night. The wakefulness regime is directly connected with the excitation of
the visual system of the brain by the light of the Sun of the visible range.
Similarly, the action of a UBF solar nature on brain biosystems at night can be
responsible for the genesis of the physiology of sleep. Earth at night
completely shields the effect on the biosphere of electromagnetic radiation,
but is transparent to solar and cosmic neutrinos [10, 11]. In view of this, and the chirality of the neutrino, it can be accepted as
the UBF. The physics of elementary particles [12] allows the decay of the solar neutrino in the interplanetary space to the
isomorphic energy forms (ν / g-EF),
consisting of the ν-shell and the
g-orbitals (Fig. 2).
A key feature of
sleep state physics is the biosynthesis in the epiphysis of the hormone melatonin
(ME), the maximum content of which in the blood is observed at 2-3 o'clock in
the morning [8]. The time
dependence of the synthesis of ME is regulated by the paired suprachiasmatic
nucleus (SCN), the main rhythm generator of the brain. SCNs have a neuronal
connection to the retina of the eyes and the epiphysis [13] therefore during
the day and when the eyes are illuminated, the SCN blocks the synthesis of ME,
but initiates it in the dark due to its spontaneous activity, which does not
cease even in the isolated state of the SCN. An important role in the activity
of SCN is played by neuropeptide Y, whose structure is based on the α-helix of 36 amino acids [14].
The main stages of
ME biosynthesis include the initiation of the suprachiasmatic nucleus into the
epiphysis through the nerve endings of L-noradrenaline, which triggers the
synthesis of the arylalkylamine-N-acetyltransferase (AANAT) enzyme. AAANAT
converts the L-tryptophan present in the epiphysis to serotonin, which, with
the participation of the hydroxyindole-O-methyltransferase enzyme, is converted
to ME (Fig. 2). To the peculiarities of the epiphysis physics, in addition to
the chirality of the participants in the biosynthesis of ME and the presence of
α-helices from the
sequences of chiral amino acids in the structure of enzymes, the dependence of
the yield of biosynthesis of ME on the effect on the epiphysis of the
alternating magnetic field (MF) is [15]. The limiting
stage in the biosynthesis of ME is the initiation of L-noradrenaline synthesis
of the enzyme AANAT, whose activity increases by two orders of magnitude [13] at night.
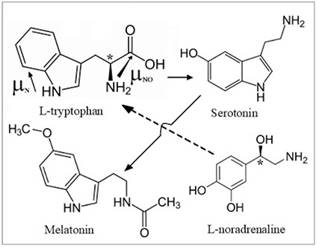
Fig. 2. Structures of the main
participants in the biosynthesis of melatonin from L-tryptophan, directions of
dipole moments of its fragments are marked by arrows (μNО ~ 10D), and asterisks are
chiral carbon atoms.
It was suggested in [11] that
solar ν/g-EFs flow around the earth
along its lithosphere (Fig.3), which up to the boundary of the Makharovichich
(h) consists of ~ 70% silica containing equal amounts of quartz L and D
crystals. On the night side of the Earth, counter flows ν/g-EF, merge and exit to the surface, affecting the biosphere. Taking into
account also the magnetosensitivity of the epiphysis, a maximum of ME content
at 2-3 nights was associated with the maximum density of the ν/g-EF effluent leaving the earth at that time. The shift of this maximum
from 0 hours to 2-3 hours was explained by the rotation of the Earth (w) and estimated the velocity (Vν) of the ν/g-EF flux across the
lithosphere as in the waveguide.
Taking into account the
Earth's rotation speed (VЕ = wRЕ) and the slope of its rotation axis (23о), the time of motion (t) of the ν/g-EF flows before their meeting at the point of the circle corresponding
to 2 o'clock in the night was expressed by the equation:
![]() ,
,
the solution of which gave Vν ~ 2.8 km/s. Knowing Vν and t, we can estimate the
displacement of the exit point of the ν/g-EF in longitude (S) relative to the diametral point of the occurrence of
the ν/g-EF flow into the earth. It
will be:
![]() ~
~
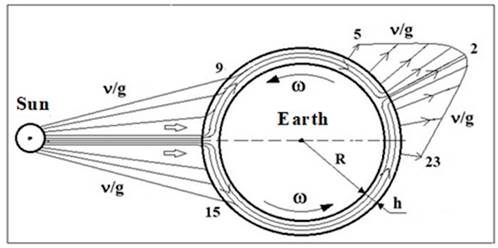
Fig. 3. Diagram of the flow of solar neutrino energy forms (ν/g) over the lithospheric layer h ~ 10-
The obtained Vν estimate of the ν/g-EF fluxes in the lithosphere is in good agreement with the magnitude of
the propagation velocity of surface seismic waves over the Earth's crust and
the probable nature of the quasiparticles (magnon and polaron) that can
participate in the mechanism of v/g-EF motion over the lithosphere [16]. The value of Vν varies depending
on the coordinates of the input of the ν/g-EF fluxes and the seasonal changes in the orientation of the Earth's
rotation axis relative to the Sun. Corresponding changes in the density of the
outgoing ν/g-EF flux affect
the metabolism of the epiphysis and the physiological parameters of the
reproductive function and psychophysics of a person that depend on it. The
chirality factor in the biochemistry of the epiphysis will be transmitted along
the neurohumoral links to other brain structures. Thus, UBF can participate in
the generation of a functional brain asymmetry resource [11].
4. Induction mechanism of saltatory conduction of nerve
fiber.
Chirality at the level of brain anatomy was manifested in
the spirals of the myelin sheaths of nerve fibers of the central nervous
system. It is not established what determines the wrapping mark of the spiral
relative to the path from the nucleus to the terminals of the neuron and what
is the distribution of the sign along the neurons of the right and left
hemispheres. It does not take into account the helicity of the myelin sheath
and in the sotator mechanism of nerve impulse transmission. These questions
remain unheeded even in theoretical works [17, 18], modeling the generation processes in the interception
of Ranvier photons with their subsequent propagation along the myelin and axon.
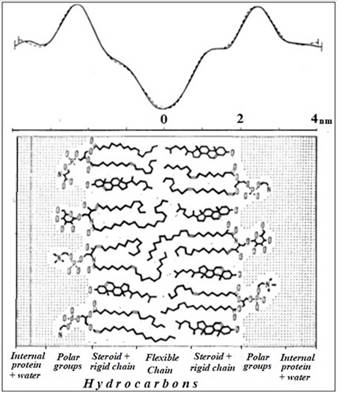
Fig. 4. Diagram of the myelin membrane structure. At the
top are the electron density profiles for the ocular and sciatic nerve, from [19].
In work [8], a mechanism was proposed for the transmission of excitation through a
myelinated fiber, based on the phenomenon of EM induction and taking into
account the chemical structure of myelin (Fig. 4). His physical essence is
explained by Fig. 5. The marginal structure of the myelin sheaths in the
intercept region of Ranvier forms spiral coils of paranodal loops of about 1 μm in length, communicating with the axoplasm through
special windows. Generation of the action potential (PD) in the Ranvier
interception is mainly due to the increase in the axoplasma concentration of Na+,
which diffuse into the paranodal region and polarize it and the axoplasm of the
paronodal loops. In the myelin sheath, the displacement current (j) is excited, the front of which moves
along the coil turns and simultaneously along the cylinder of the myelin
segment. In accordance with the Maxwell equations (1) with this displacement
current, a vortex magnetic field will be associated, whose front will move
along the axon as in the core with a velocity V ~ C/n. The sign of the myelin
helix determines the direction of rotation of the current and, according to the
rule of the right screw, the direction of the vector B (Fig. 5).
The kinetics of ion currents and displacement currents in
the axon, membrane, and paranodal loops of the myelin sheath correlates with
the kinetics of growth and subsequent relaxation of the membrane potential.
Since the phase of the PD growth lasts about 0.1 - 0.2 ms, and the relaxation
time of the membrane potential is of the order of 1 ms [8], then the displacement currents
corresponding to the phase of increase will be an order of magnitude greater
than the relaxation currents. Thus, the generation of PDs in the Ranvier
interception is associated with the induction in the nerve fiber of vortex EM
quanta, similar to the ν/g-EF in Fig.
1. It is possible that this is the main function of the end coils of myelin
sheaths and incision spirals.
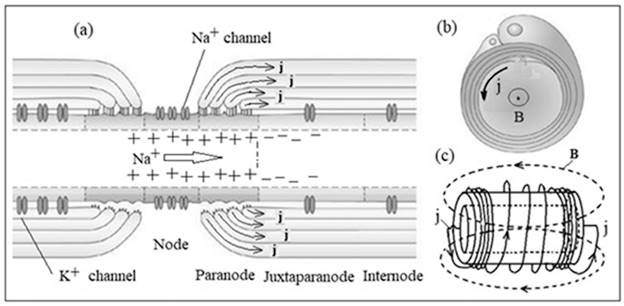
Fig. 5. Induction model of the
sotator mechanism of nerve impulse transmission. Scheme of interception of
Ranvier (a), myelin sheath (b) and myelin nerve segment (c). The arrow shows
the diffusion of Na+, j -
the displacement currents, B - the vortex magnetic field. The original figures
(a) and (b) are taken from [18].
The direction of the vector of the flux density of the EM
energy (the Poiting vector) will be determined by the sign of the spiral. This
chirality factor of the neuron will ensure one-sided distribution of the
EM-quantum and PD on the myelinated nerve. When the EM-quantum of the terminal
coil of the myelin segment is reached, it will play the role of a stimulus for
the generation of PD in the next interception of Ranvier.
This reaction is mainly due to sodium channels due to the
fact that the magnetic moment of Na+ is 6 times, and the magnetic
susceptibility is 200 times larger than the corresponding values for
K+ [8]. In the inductive model of the saltator conduction of the
neuron, the speed of the spike motion will be limited by the process of current
excitation in the end coils, whose time is of the order of 10-6 s (1
μm:1 m/s). At
the same time, the average transfer rate of PD from one end of the myelin
segment to the other at a length of about 100 μm and will determine the rate of the saltative mechanism
of conductivity ~ 100 m/s.
The inductive mechanism of the
saltatory conduction of the nerve can be taken into account in the equivalent
electrical circuit of the membrane [8], simulating the spiral myelin sheath
with an inductor (Fig. 6). The transformation of the circuit into a circuit of
parallel connected oscillatory circuits substantially extends the range of
modeling the electrophysical properties of the neuron.
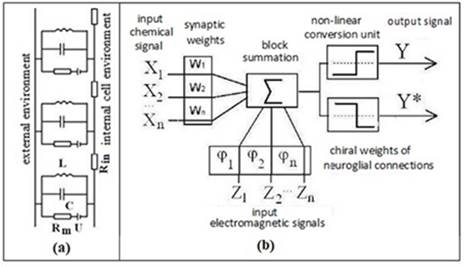
Fig. 6. (a) Modified electrical
circuit of the nerve membrane. Rm, U - ion channel; C is the
capacity of the membrane; L - inductance of glial membrane myelin helices; Rin
- resistance of axoplasm. (b) Modified functional scheme of a formal neuron
[21]. Xn - biochemical, Zn - electrophysical factors of
neuron activity; Y («yes»), Y* («no») are analogues of the exciting and braking
signals.
The element of inductance in
the electrical circuit of the nerve membrane also makes it possible to model
the chirality factor of the neuron and to associate it with the mechanism of
the differentiation of nerve signals to stimulating and inhibitory ones. The
combination of the chirality factor with the biochemical factor (synaptic
connections) empowers the logical element of neural networks to encode the
«yes» and «no» signals (Fig. 6).
5. Physics of integrative mechanisms of the brain.
Interactions of EF with each
other and with the material of the brain obey the isoenergetic
fractal-resonance rearrangements of the structures themselves of the EF and
cooperative ensembles from the dynamic molecular-cellular elements of the brain
[6-9]. The liquid media of the visual and auditory systems of the brain,
as well as its cerebrospinal and circulatory system, which, in principle,
possess chirality should be referred to the latter as the first [22]. The
mechanism of heuristic thinking integrates the functions of individual organs
responsible for the following processes: perception of internal and external
signals, their processing, synthesis at the level of new patterns, recognition
of novelty of information, evaluation of its significance and memorization.
With each stage of the
mechanism of thinking, a definite dominant in the frequency spectrum of
electrical oscillators of the heart and brain, extending from ~0.5 to ~100 Hz,
will be associated [8, 9]. The rhythmic dynamics of these processes at the
EM-level of the entire brain can be modeled by a chiral integral EM-scheme
(Fig. 7). The common mechanism of excitation of internal rhythms of the brain
is the phenomenon of EM-induction and the interaction of vibrational
EM-circuits between themselves. For example, the alpha rhythm of the background
electrical activity of the cortex maintains at a proper level the stability of
the neocortex and thalamus connections. Right-left parts of the thalamus and
the cortex of the hemispheres can be represented by dissimilar plates of two
spherical capacitors, and the nerve connections between them (the radiance of
the thalamus) will simulate ohmic connections and inductive coils in equivalent
schemes of circuits operating at a frequency of alpha rhythm. The asymmetry of
the inductive elements of the EM contours of the right and left hemispheres can
underlie the specialization of their cognitive functions.
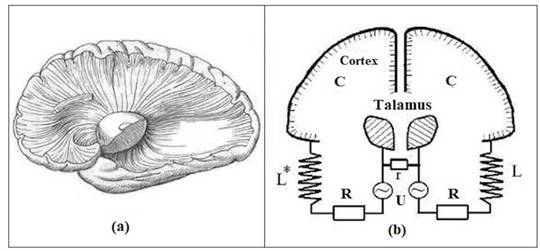
Fig. 7. (a) Radiation of the
thalamus. (b) Equivalent oscillatory circuits modeling the alpha rhythms of the
brain. L, L*, R - inductive and ohmic models of the radiance of the thalamus
(sign *) means a mirror inversion of the chiral structures of the right
hemisphere); r - intertalamic fusion; C and U are the capacitance and potential
difference between the thalamus and the cortex.
6. Conclusion.
The evolution of natural
science already has one dialectical leap in the form of a transition from
classical physics to quantum physics. In the present work it was shown that in
order to understand the mechanism of human heuristic thinking, it is necessary
to deepen the physics of the brain to an elementary level of organization of
matter in Maxwell and to take into account the chirality of external and
internal morphofunctional factors. In parallel with this, fundamental physics
will solve the neutrino problem, which physicists call the «window into a new
physics» [23]. The discovery of this «window» may be
explaining the key role of solar neutrino physics in the evolution of a homo
sapiens.
References
1. Holmansky AS, Minakhin AA Factors of phylogenesis of human posture and
morphogenesis. Sciences. 2012. 4. 1-8.
http://naukovedenie.ru/PDF/48pvn412.pdf
2. Maxwell, J. Selected papers on the theory of the electromagnetic field.
M. 1954
3. Kaplan A. Ya. Harmony of the Big Bang. Domestic notes. 2014.2.123-36.
4. Khoruzhiy S. The discourses of the inner and the outer in the practices
of oneself. Moscow Psychotherapeutic
Journal. 2003. 3. 5-25
5. Kizel V.A. Physical causes of dissymmetry of living systems. M.1985.
120.
6. Kholmanskiy A.S. Chirality and quantum effects as factors of
morphogenesis. Mathematical morphology:
electronic mathematical and medico-biological journal. 2010. 9. http://sgma.alpha-design.ru/MMORPH/N-28-html/kholmanskiy-2/kholmanskiy-2.htm
7. Kholmanskiy A.S. Elementary physics of ether. Science and Peace. 2016. 1
(4).19.
http://scienceph.ru/d/413259/d/science_and_world_no_4_(32),_april,_vol._i.pdf
8. Kholmanskiy A. Modeling of brain physics. Mathematical morphology. Electronic mathematical and Medico-biological
journal. 2006. 5 (4); http://new-idea.kulichki.net/pubfiles/180520162644.pdf
9. Kholmanskiy AS, Minakhin AA Interconnection of electrical oscillations
of the heart and brain. Bulletin of St.
Petersburg State University. Medicine. 2018. 13 (2).
10. Kholmanskiy A.S. Galactic factor of spiritual evolution. Asymmetry. 2009. 3 (1). 47. http://cerebral-asymmetry.narod.ru/Asymmetry_1_2009.pdf
11. Kholmanskiy A.S. Dependence of the resource of functional asymmetry of
the brain on external conditions. Ibid.
2009. 3 (1). 51-6: http://cerebral-asymmetry.narod.ru/Asymmetry_1_2009.pdf .
12. Klapdor-Kleingrothaus G.V., Staudt A. Non-accelerating physics of
elementary particles. M. 1997.528.
13. Kovalzon VM Foundations of somnology: physiology and neurochemistry of
the cycle «wakefulness-sleep». M. 2014.239.
14. Medanic M., Gillette M.U. Suprachiasmatic circadian pacemaker of the
rat shows two windows of sensitivity to neuropeptide Y in vitro. Brain Research, 1993. 620 (2), 281-6.
15. Temur'yants N., Shekhotkin A., Magnitosensitivity of the epiphysis. Biophysics. 1998. 43. 761.
16. Aleshkevich VA, Dedenko LG, Karavaev VA Fluctuations and waves. Moscow
State University, 2001.
17. Kumar S. et al. Possible existence of optical communication channels in
the brain. Sci Rep. 2016; 6: 36508. doi:
10.1038 / srep36508
18. Zangari A., Micheli D., Galeazzi R., Tozzi A. Node of Ranvier as an
Array of Bio-Nanoantennas for Infrared Communication in Nerve Tissue. Scientific Repots. 2018. 8. 539.
19. Volkenshtein M.V. Biophysics: M. 1988.592
20. Poliak S., Peles E. The local differentiation of myelinated axons at
the nodes of Ranvier. Nature Rev.
Neuroscience. 2003. 4. 12.
968-80
21. Terekhov S.A. Lectures on the theory and applications of artificial
neural networks. Snezhinsk, 1994-1998.
22. Kholmanskiy A. Chirality anomalies of water solutions of saccharides. J. Mol. Liq. 2016.216. 683.
23. Giunti C., Studenikin A. Neutrino electromagnetic interactions: a window to new physics. Rev. Mod. Phys. 2015. 87.531
aThe Scientific Center «Bemkom», 11, Shenkursky proezd, Moscow, 127340, Russian Federation
Поступила в редакцию 22.05.2018